 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5mps | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
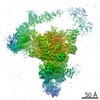
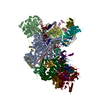
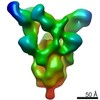
| タイトル | Structure of a spliceosome remodeled for exon ligation | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | SPLICING / pre-mRNA splicing / trans-esterification / lariat intermediate / complex C-star | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報U2-type post-spliceosomal complex / mRNA branch site recognition / U2-type post-mRNA release spliceosomal complex / cellular bud site selection / pre-mRNA 3'-splice site binding / post-mRNA release spliceosomal complex / nuclear mRNA surveillance / generation of catalytic spliceosome for first transesterification step / cis assembly of pre-catalytic spliceosome / spliceosome conformational change to release U4 (or U4atac) and U1 (or U11) ...U2-type post-spliceosomal complex / mRNA branch site recognition / U2-type post-mRNA release spliceosomal complex / cellular bud site selection / pre-mRNA 3'-splice site binding / post-mRNA release spliceosomal complex / nuclear mRNA surveillance / generation of catalytic spliceosome for first transesterification step / cis assembly of pre-catalytic spliceosome / spliceosome conformational change to release U4 (or U4atac) and U1 (or U11) / U4/U6 snRNP / 7-methylguanosine cap hypermethylation / pICln-Sm protein complex / U2-type catalytic step 1 spliceosome / pre-mRNA binding / snRNP binding / small nuclear ribonucleoprotein complex / splicing factor binding / SMN-Sm protein complex / spliceosomal tri-snRNP complex / commitment complex / mRNA cis splicing, via spliceosome / U2-type spliceosomal complex / U2-type prespliceosome assembly / U2-type catalytic step 2 spliceosome / U4 snRNP / U2 snRNP / U1 snRNP / poly(U) RNA binding / U2-type prespliceosome / precatalytic spliceosome / generation of catalytic spliceosome for second transesterification step / Formation of TC-NER Pre-Incision Complex / spliceosomal complex assembly / mRNA 5'-splice site recognition / mRNA 3'-splice site recognition / Gap-filling DNA repair synthesis and ligation in TC-NER / DNA replication origin binding / spliceosomal tri-snRNP complex assembly / Dual incision in TC-NER / Prp19 complex / U5 snRNA binding / U5 snRNP / DNA replication initiation / U2 snRNA binding / U6 snRNA binding / pre-mRNA intronic binding / spliceosomal snRNP assembly / U1 snRNA binding / U4/U6 x U5 tri-snRNP complex / positive regulation of cell cycle / catalytic step 2 spliceosome / nuclear periphery / positive regulation of RNA splicing / spliceosomal complex / mRNA splicing, via spliceosome / metallopeptidase activity / GTPase activity / mRNA binding / chromatin binding / chromatin / GTP binding / DNA binding / RNA binding / zinc ion binding / nucleus / cytoplasm / cytosol 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.85 Å | ||||||||||||
 データ登録者 データ登録者 | Fica, S.M. / Oubridge, C. / Galej, W.P. / Wilkinson, M.E. / Newman, A.J. / Bai, X.-C. / Nagai, K. | ||||||||||||
| 資金援助 |  英国, 3件 英国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2017 タイトル: Structure of a spliceosome remodelled for exon ligation. 著者: Sebastian M Fica / Chris Oubridge / Wojciech P Galej / Max E Wilkinson / Xiao-Chen Bai / Andrew J Newman / Kiyoshi Nagai / 要旨: The spliceosome excises introns from pre-mRNAs in two sequential transesterifications-branching and exon ligation-catalysed at a single catalytic metal site in U6 small nuclear RNA (snRNA). Recently ...The spliceosome excises introns from pre-mRNAs in two sequential transesterifications-branching and exon ligation-catalysed at a single catalytic metal site in U6 small nuclear RNA (snRNA). Recently reported structures of the spliceosomal C complex with the cleaved 5' exon and lariat-3'-exon bound to the catalytic centre revealed that branching-specific factors such as Cwc25 lock the branch helix into position for nucleophilic attack of the branch adenosine at the 5' splice site. Furthermore, the ATPase Prp16 is positioned to bind and translocate the intron downstream of the branch point to destabilize branching-specific factors and release the branch helix from the active site. Here we present, at 3.8 Å resolution, the cryo-electron microscopy structure of a Saccharomyces cerevisiae spliceosome stalled after Prp16-mediated remodelling but before exon ligation. While the U6 snRNA catalytic core remains firmly held in the active site cavity of Prp8 by proteins common to both steps, the branch helix has rotated by 75° compared to the C complex and is stabilized in a new position by Prp17, Cef1 and the reoriented Prp8 RNase H-like domain. This rotation of the branch helix removes the branch adenosine from the catalytic core, creates a space for 3' exon docking, and restructures the pairing of the 5' splice site with the U6 snRNA ACAGAGA region. Slu7 and Prp18, which promote exon ligation, bind together to the Prp8 RNase H-like domain. The ATPase Prp22, bound to Prp8 in place of Prp16, could interact with the 3' exon, suggesting a possible basis for mRNA release after exon ligation. Together with the structure of the C complex, our structure of the C* complex reveals the two major conformations of the spliceosome during the catalytic stages of splicing. #1: ジャーナル: Science / 年: 2015 タイトル: Structural basis of pre-mRNA splicing. 著者: Jing Hang / Ruixue Wan / Chuangye Yan / Yigong Shi /  要旨: Splicing of precursor messenger RNA is performed by the spliceosome. In the cryogenic electron microscopy structure of the yeast spliceosome, U5 small nuclear ribonucleoprotein acts as a central ...Splicing of precursor messenger RNA is performed by the spliceosome. In the cryogenic electron microscopy structure of the yeast spliceosome, U5 small nuclear ribonucleoprotein acts as a central scaffold onto which U6 and U2 small nuclear RNAs (snRNAs) are intertwined to form a catalytic center next to Loop I of U5 snRNA. Magnesium ions are coordinated by conserved nucleotides in U6 snRNA. The intron lariat is held in place through base-pairing interactions with both U2 and U6 snRNAs, leaving the variable-length middle portion on the solvent-accessible surface of the catalytic center. The protein components of the spliceosome anchor both 5' and 3' ends of the U2 and U6 snRNAs away from the active site, direct the RNA sequences, and allow sufficient flexibility between the ends and the catalytic center. Thus, the spliceosome is in essence a protein-directed ribozyme, with the protein components essential for the delivery of critical RNA molecules into close proximity of one another at the right time for the splicing reaction. #2: ジャーナル: Nature / 年: 2015タイトル: The architecture of the spliceosomal U4/U6.U5 tri-snRNP. 著者: Thi Hoang Duong Nguyen / Wojciech P Galej / Xiao-chen Bai / Christos G Savva / Andrew J Newman / Sjors H W Scheres / Kiyoshi Nagai / 要旨: U4/U6.U5 tri-snRNP is a 1.5-megadalton pre-assembled spliceosomal complex comprising U5 small nuclear RNA (snRNA), extensively base-paired U4/U6 snRNAs and more than 30 proteins, including the key ...U4/U6.U5 tri-snRNP is a 1.5-megadalton pre-assembled spliceosomal complex comprising U5 small nuclear RNA (snRNA), extensively base-paired U4/U6 snRNAs and more than 30 proteins, including the key components Prp8, Brr2 and Snu114. The tri-snRNP combines with a precursor messenger RNA substrate bound to U1 and U2 small nuclear ribonucleoprotein particles (snRNPs), and transforms into a catalytically active spliceosome after extensive compositional and conformational changes triggered by unwinding of the U4 and U6 (U4/U6) snRNAs. Here we use cryo-electron microscopy single-particle reconstruction of Saccharomyces cerevisiae tri-snRNP at 5.9 Å resolution to reveal the essentially complete organization of its RNA and protein components. The single-stranded region of U4 snRNA between its 3' stem-loop and the U4/U6 snRNA stem I is loaded into the Brr2 helicase active site ready for unwinding. Snu114 and the amino-terminal domain of Prp8 position U5 snRNA to insert its loop I, which aligns the exons for splicing, into the Prp8 active site cavity. The structure provides crucial insights into the activation process and the active site of the spliceosome. #3: ジャーナル: Nature / 年: 2016タイトル: Cryo-EM structure of the yeast U4/U6.U5 tri-snRNP at 3.7 Å resolution. 著者: Thi Hoang Duong Nguyen / Wojciech P Galej / Xiao-Chen Bai / Chris Oubridge / Andrew J Newman / Sjors H W Scheres / Kiyoshi Nagai / 要旨: U4/U6.U5 tri-snRNP represents a substantial part of the spliceosome before activation. A cryo-electron microscopy structure of Saccharomyces cerevisiae U4/U6.U5 tri-snRNP at 3.7 Å resolution led ...U4/U6.U5 tri-snRNP represents a substantial part of the spliceosome before activation. A cryo-electron microscopy structure of Saccharomyces cerevisiae U4/U6.U5 tri-snRNP at 3.7 Å resolution led to an essentially complete atomic model comprising 30 proteins plus U4/U6 and U5 small nuclear RNAs (snRNAs). The structure reveals striking interweaving interactions of the protein and RNA components, including extended polypeptides penetrating into subunit interfaces. The invariant ACAGAGA sequence of U6 snRNA, which base-pairs with the 5'-splice site during catalytic activation, forms a hairpin stabilized by Dib1 and Prp8 while the adjacent nucleotides interact with the exon binding loop 1 of U5 snRNA. Snu114 harbours GTP, but its putative catalytic histidine is held away from the γ-phosphate by hydrogen bonding to a tyrosine in the amino-terminal domain of Prp8. Mutation of this histidine to alanine has no detectable effect on yeast growth. The structure provides important new insights into the spliceosome activation process leading to the formation of the catalytic centre. #4: ジャーナル: Nature / 年: 2016タイトル: Cryo-EM structure of the spliceosome immediately after branching. 著者: Wojciech P Galej / Max E Wilkinson / Sebastian M Fica / Chris Oubridge / Andrew J Newman / Kiyoshi Nagai / 要旨: Precursor mRNA (pre-mRNA) splicing proceeds by two consecutive transesterification reactions via a lariat-intron intermediate. Here we present the 3.8 Å cryo-electron microscopy structure of the ...Precursor mRNA (pre-mRNA) splicing proceeds by two consecutive transesterification reactions via a lariat-intron intermediate. Here we present the 3.8 Å cryo-electron microscopy structure of the spliceosome immediately after lariat formation. The 5'-splice site is cleaved but remains close to the catalytic Mg site in the U2/U6 small nuclear RNA (snRNA) triplex, and the 5'-phosphate of the intron nucleotide G(+1) is linked to the branch adenosine 2'OH. The 5'-exon is held between the Prp8 amino-terminal and linker domains, and base-pairs with U5 snRNA loop 1. Non-Watson-Crick interactions between the branch helix and 5'-splice site dock the branch adenosine into the active site, while intron nucleotides +3 to +6 base-pair with the U6 snRNA ACAGAGA sequence. Isy1 and the step-one factors Yju2 and Cwc25 stabilize docking of the branch helix. The intron downstream of the branch site emerges between the Prp8 reverse transcriptase and linker domains and extends towards the Prp16 helicase, suggesting a plausible mechanism of remodelling before exon ligation. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5mps.cif.gz | 1.6 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5mps.ent.gz | 1.2 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5mps.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5mps_validation.pdf.gz | 1.5 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5mps_full_validation.pdf.gz | 1.6 MB | 表示 | |
| XML形式データ | 5mps_validation.xml.gz | 175.5 KB | 表示 | |
| CIF形式データ | 5mps_validation.cif.gz | 286.5 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/mp/5mpsftp://data.pdbj.org/pub/pdb/validation_reports/mp/5mps | HTTPS FTP |
-関連構造データ
| 関連構造データ |  3539MC  3541C  3542C  5mq0C M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ | |
| 電子顕微鏡画像生データ | EMPIAR-10687 (タイトル: Yeast C, Ci, C*, and P complex spliceosomes / Data size: 8.9 TB Data #1: Unaligned movies of C-complex spliceosome with 3' splice site AG to AC mutation (Dataset 1) [micrographs - multiframe] Data #2: Unaligned movies of C and C*-complex spliceosomes with 3' splice site AG to AdG mutation (Dataset 2) [micrographs - multiframe] Data #3: Unaligned movies of C and C*-complex spliceosomes with 3' splice site AG to AdG mutation (Dataset 3) [micrographs - multiframe] Data #4: Aligned movies of C-complex spliceosomes with cold-sensitive prp16-302 mutation, purified with Cwc25 (Dataset 4) [micrographs - multiframe] Data #5: Unaligned movies of C-complex spliceosomes with cold-sensitive prp16-302 mutation, purified with Cwc25 and incubated with ATP and Mg (Dataset 5) [micrographs - multiframe] Data #6: Unaligned movies of C, C*, and P-complex spliceosomes with dominant-negative Prp22 mutation K512A, purified with Slu7 (Dataset 6) [micrographs - multiframe] Data #7: Unaligned movies of P-complex spliceosomes with dominant-negative Prp22 mutation K512A, treated with anti-3'exon RNaseH oligo, purified in presence of Mg (Dataset 9) [micrographs - single frame] Data #8: Selected C-complex particles after polishing [picked particles - single frame - processed] Data #9: Selected P-complex particles after polishing [picked particles - single frame - processed] Data #10: Various signal subtractions for C- and P-complex spliceosomes [picked particles - single frame - processed]) |




-リンク
 PDBj
PDBj


































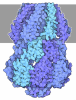

- 集合体
集合体
| 登録構造単位 | 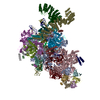
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 5種, 5分子 IE265
| #1: RNA鎖 | 分子量: 30200.730 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: Saccharomyces cerevisiae UBC4 pre-mRNA intron, in vitro transcribed. 由来: (組換発現) 発現宿主: in vitro transcription vector pT7-Fluc(deltai) (その他) 参照: GenBank: 4718 |
|---|---|
| #2: RNA鎖 | 分子量: 6518.976 Da / 分子数: 1 / 由来タイプ: 組換発現 詳細: Saccharomyces cerevisiae UBC4 pre-mRNA exon, in vitro transcribed. 由来: (組換発現) 遺伝子: UBC4 発現宿主: in vitro transcription vector pT7-Fluc(deltai) (その他) |
| #3: RNA鎖 | 分子量: 376267.406 Da / 分子数: 1 / 由来タイプ: 組換発現 / 詳細: Saccharomyces cerevisiae U2 snRNA 由来: (組換発現) 発現宿主: |
| #4: RNA鎖 | 分子量: 35883.176 Da / 分子数: 1 / 由来タイプ: 組換発現 / 詳細: Saccharomyces cerevisiae U6 snRNA 由来: (組換発現) 発現宿主: |
| #5: RNA鎖 | 分子量: 57444.875 Da / 分子数: 1 / 由来タイプ: 組換発現 / 詳細: Saccharomyces cerevisiae U5 snRNA 由来: (組換発現) 発現宿主: |
-Pre-mRNA-splicing factor ... , 15種, 15分子 ACHJLMNOPRSTacy
| #6: タンパク質 | 分子量: 279867.469 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Prp8 protein / 由来: (天然) |
|---|---|
| #7: タンパク質 | 分子量: 114174.008 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: c Snu114 protein / 由来: (天然) |
| #8: タンパク質 | 分子量: 67386.062 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Cwc22 protein / 由来: (天然) |
| #9: タンパク質 | 分子量: 50771.289 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Prp46 protein / 由来: (天然) |
| #11: タンパク質 | 分子量: 18484.502 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Bud31 protein / 由来: (天然) |
| #12: タンパク質 | 分子量: 38486.562 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Cwc2 protein / 由来: (天然) |
| #13: タンパク質 | 分子量: 40988.590 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Ecm2 protein / 由来: (天然) |
| #14: タンパク質 | 分子量: 67837.773 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Cef1 protein / 由来: (天然) |
| #15: タンパク質 | 分子量: 19975.195 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Cwc15 protein / 由来: (天然) |
| #16: タンパク質 | 分子量: 15793.596 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Cwc21 protein / 由来: (天然) |
| #17: タンパク質 | 分子量: 82555.859 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Clf1 protein / 由来: (天然) |
| #18: タンパク質 | 分子量: 101875.852 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Syf1 protein / 由来: (天然) |
| #19: タンパク質 | 分子量: 28414.391 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Prp18 protein / 由来: (天然) |
| #20: タンパク質 | 分子量: 44722.875 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Slu7 protein / 由来: (天然) |
| #23: タンパク質 | 分子量: 24850.719 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Syf2 protein / 由来: (天然) |
-Pre-mRNA-processing ... , 2種, 2分子 Ko
| #10: タンパク質 | 分子量: 42548.727 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Prp45 protein / 由来: (天然) |
|---|---|
| #21: タンパク質 | 分子量: 52128.762 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae Prp17 protein / 由来: (天然) |
-タンパク質 , 2種, 2分子 Xb
| #22: タンパク質 | 分子量: 5805.147 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Unknown protein helices within C-star complex / 由来: (天然) |
|---|---|
| #24: タンパク質 | 分子量: 22426.990 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae SmB protein / 由来: (天然) |
-Small nuclear ribonucleoprotein ... , 6種, 6分子 defghj
| #25: タンパク質 | 分子量: 11240.139 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae SmD3 protein / 由来: (天然) |
|---|---|
| #26: タンパク質 | 分子量: 10385.098 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae SmE protein / 由来: (天然) |
| #27: タンパク質 | 分子量: 9669.945 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae SmF protein / 由来: (天然) |
| #28: タンパク質 | 分子量: 8490.809 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae SmG protein / 由来: (天然) |
| #29: タンパク質 | 分子量: 16296.798 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae SmD1 protein / 由来: (天然) |
| #30: タンパク質 | 分子量: 12876.066 Da / 分子数: 1 / 由来タイプ: 天然 / 詳細: Saccharomyces cerevisiae SmD2 protein / 由来: (天然) |
-非ポリマー , 5種, 13分子 

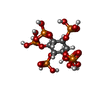
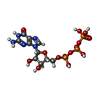

| #31: 化合物 |  分子量: 24.305 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Mg#32: 化合物 |  分子量: 39.098 Da / 分子数: 2 / 由来タイプ: 合成 / 式: K 分子量: 39.098 Da / 分子数: 2 / 由来タイプ: 合成 / 式: K#33: 化合物 | ChemComp-IHP / |  分子量: 660.035 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C6H18O24P6 分子量: 660.035 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C6H18O24P6#34: 化合物 | ChemComp-GTP / |  分子量: 523.180 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H16N5O14P3 / コメント: GTP, エネルギー貯蔵分子*YM 分子量: 523.180 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C10H16N5O14P3 / コメント: GTP, エネルギー貯蔵分子*YM#35: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 6 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 6 / 由来タイプ: 合成 / 式: Zn |
|---|
-詳細
| Has protein modification | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Saccharomyces cerevisiae spliceosome. Complex C just after Prp16-mediated remodeling タイプ: COMPLEX 詳細: Splicing extract was prepared from Slu7-TAPS yeast strains. An in vitro transcribed yeast UBC4 pre-mRNA substrate (with 2 x MS2 bacteriophage coat protein-binding stem loops at the 5' end and ...詳細: Splicing extract was prepared from Slu7-TAPS yeast strains. An in vitro transcribed yeast UBC4 pre-mRNA substrate (with 2 x MS2 bacteriophage coat protein-binding stem loops at the 5' end and with a 2'-deoxy substitution at the 3'-splice site sequence UAG sequence (UA-2'dG) was pre-bound to an MS2-maltose binding protein fusion protein. This substrate-protein complex was added to the splicing extract. The splicing reaction proceeded through the first step but the second step was blocked by the deoxy substitution. Substrate-bound spliceosomes from the splicing extract were purified on amylose resin and eluted with maltose. Subsequently the spliceosomes were captured on streptactin resin and eluted with desthiobiotin. Purified spliceosomes were concentrated in 20 mM HEPES KOH pH 7.9, 100 mM KCl, 0.25 mM EDTA. Entity ID: #1-#30 / 由来: MULTIPLE SOURCES | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | pH: 7.9 / 詳細: NP-40 is also called IGEPAL CA-630 | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.3 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 400 divisions/in. / グリッドのタイプ: Quantifoil R1.2/1.3 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK III / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K 詳細: 3.5 microlitres sample were applied to the grid, left for 25 seconds and then blotted for 3.0-3.5 seconds before plunging. |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 詳細: GIF Quantum energy filter, 20 eV slit width |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 4500 nm / 最小 デフォーカス(公称値): 500 nm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 0.8 sec. / 電子線照射量: 2 e/Å2 / 検出モード: SUPER-RESOLUTION フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 実像数: 3596 詳細: Total dose: 40 electrons/Angstrom^2 over 16 seconds. 20 movie frames collected at 1.25 frames per second. |
| 電子光学装置 | エネルギーフィルター名称: GIF Quantum |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0124 / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 350000 詳細: Selected initial particles automatically using C complex 2D class averages, low-pass filtered to 20 Angstrom for automatic particle picking. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.85 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 65824 / クラス平均像の数: 4 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | B value: 330 / プロトコル: FLEXIBLE FIT / 空間: RECIPROCAL / Target criteria: Fourier Shell Correlation 詳細: Used secondary structure restraints generated in ProSMART and LibG. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 3.85→257.4 Å / Cor.coef. Fo:Fc: 0.952 / SU B: 37.73 / SU ML: 0.526 / ESU R: 0.868 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 溶媒モデル: PARAMETERS FOR MASK CACLULATION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 170.644 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 合計: 59157 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|