 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5m1j | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Nonstop ribosomal complex bound with Dom34 and Hbs1 | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | RIBOSOME / mRNA surveillance | |||||||||
| Function / homology |  Function and homology information Function and homology informationEukaryotic Translation Elongation / Dom34-Hbs1 complex / RNA surveillance / nuclear-transcribed mRNA catabolic process, no-go decay / nuclear-transcribed mRNA catabolic process, non-stop decay / HSF1 activation / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, LSU-rRNA,5S) / regulation of amino acid metabolic process / negative regulation of glucose mediated signaling pathway / translational readthrough ...Eukaryotic Translation Elongation / Dom34-Hbs1 complex / RNA surveillance / nuclear-transcribed mRNA catabolic process, no-go decay / nuclear-transcribed mRNA catabolic process, non-stop decay / HSF1 activation / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, LSU-rRNA,5S) / regulation of amino acid metabolic process / negative regulation of glucose mediated signaling pathway / translational readthrough / : / positive regulation of translational fidelity / RMTs methylate histone arginines / Protein methylation / mTORC1-mediated signalling / Protein hydroxylation / ribosome disassembly / positive regulation of protein kinase activity / ribosome-associated ubiquitin-dependent protein catabolic process / pre-mRNA 5'-splice site binding / GDP-dissociation inhibitor activity / cytosolic large ribosomal subunit assembly / nonfunctional rRNA decay / Formation of the ternary complex, and subsequently, the 43S complex / Translation initiation complex formation / positive regulation of nuclear-transcribed mRNA catabolic process, deadenylation-dependent decay / Ribosomal scanning and start codon recognition / response to cycloheximide / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / preribosome, small subunit precursor / Major pathway of rRNA processing in the nucleolus and cytosol / mRNA destabilization / SRP-dependent cotranslational protein targeting to membrane / GTP hydrolysis and joining of the 60S ribosomal subunit / negative regulation of mRNA splicing, via spliceosome / preribosome, large subunit precursor / Formation of a pool of free 40S subunits / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / L13a-mediated translational silencing of Ceruloplasmin expression / negative regulation of translational frameshifting / translational elongation / positive regulation of translational initiation / ribosomal large subunit export from nucleus / Ub-specific processing proteases / G-protein alpha-subunit binding / 90S preribosome / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translation elongation factor activity / ribosomal subunit export from nucleus / translational termination / regulation of translational fidelity / maturation of LSU-rRNA / protein-RNA complex assembly / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / Neutrophil degranulation / translation regulator activity / RNA endonuclease activity / ribosomal small subunit export from nucleus / DNA-(apurinic or apyrimidinic site) endonuclease activity / rescue of stalled cytosolic ribosome / cellular response to amino acid starvation / protein kinase C binding / ribosomal large subunit biogenesis / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / ribosome assembly / meiotic cell cycle / positive regulation of translation / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / macroautophagy / maturation of SSU-rRNA / translational initiation / small-subunit processome / maintenance of translational fidelity / modification-dependent protein catabolic process / cytoplasmic stress granule / protein tag activity / rRNA processing / ribosomal small subunit assembly / ribosome biogenesis / ribosome binding / ribosomal small subunit biogenesis / 5S rRNA binding / ribosomal large subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / Hydrolases; Acting on acid anhydrides; Acting on GTP to facilitate cellular and subcellular movement / cytoplasmic translation / negative regulation of translation / rRNA binding / structural constituent of ribosome / protein ubiquitination / ribosome / translation / G protein-coupled receptor signaling pathway / negative regulation of gene expression / cell division Similarity search - Function | |||||||||
| Biological species |  synthetic construct (others) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.3 Å | |||||||||
 Authors Authors | Hilal, T. / Yamamoto, H. / Loerke, J. / Buerger, J. / Mielke, T. / Spahn, C.M.T. | |||||||||
| Funding support |  Germany, 2items Germany, 2items
| |||||||||
 Citation Citation | Journal: Nat Commun / Year: 2016 Title: Structural insights into ribosomal rescue by Dom34 and Hbs1 at near-atomic resolution. Authors: Tarek Hilal / Hiroshi Yamamoto / Justus Loerke / Jörg Bürger / Thorsten Mielke / Christian M T Spahn / Abstract: The surveillance of mRNA translation is imperative for homeostasis. Monitoring the integrity of the message is essential, as the translation of aberrant mRNAs leads to stalling of the translational ...The surveillance of mRNA translation is imperative for homeostasis. Monitoring the integrity of the message is essential, as the translation of aberrant mRNAs leads to stalling of the translational machinery. During ribosomal rescue, arrested ribosomes are specifically recognized by the conserved eukaryotic proteins Dom34 and Hbs1, to initiate their recycling. Here we solve the structure of Dom34 and Hbs1 bound to a yeast ribosome programmed with a nonstop mRNA at 3.3 Å resolution using cryo-electron microscopy. The structure shows that Domain N of Dom34 is inserted into the upstream mRNA-binding groove via direct stacking interactions with conserved nucleotides of 18S rRNA. It senses the absence of mRNA at the A-site and part of the mRNA entry channel by direct competition. Thus, our analysis establishes the structural foundation for the recognition of aberrantly stalled 80S ribosomes by the Dom34·Hbs1·GTP complex during Dom34-mediated mRNA surveillance pathways. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5m1j.cif.gz | 4.9 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5m1j.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 5m1j.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/m1/5m1jftp://data.pdbj.org/pub/pdb/validation_reports/m1/5m1j | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4140MC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |
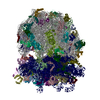
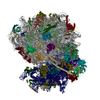
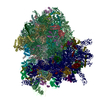
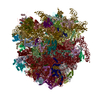

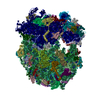
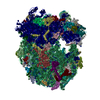
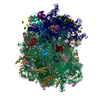
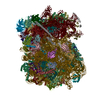
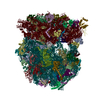
-Links
 PDBj
PDBj


































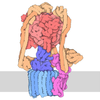




- Assembly
Assembly
| Deposited unit | 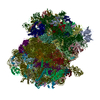
|
|---|---|
| 1 |
|
-Components
-Protein , 5 types, 5 molecules A1g2f2m5A6
| #1: Protein | Mass: 43574.344 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: Hexahistidine-tagged protein Source: (gene. exp.) Gene: DOM34, YNL001W, N2016 / Production host:  References: UniProt: P33309, Hydrolases; Acting on ester bonds |
|---|---|
| #14: Protein | Mass: 34710.023 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #35: Protein | Mass: 8101.675 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #61: Protein | Mass: 6032.321 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #80: Protein | Mass: 68812.383 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Details: Hexahistidine tagged protein Source: (gene. exp.) Gene: HBS1, YKR084C, YKR404 / Production host: |
+40S ribosomal protein ... , 31 types, 31 molecules A2a2B2b2C2c2D2d2E2e2F2G2H2I2J2K2L2M2N2O2P2Q2R2S2T2U2V2W2X2Y2Z2
-RNA chain , 6 types, 6 molecules 22143444X7A3
| #34: RNA chain | Mass: 579126.562 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #36: RNA chain | Mass: 1097493.875 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #37: RNA chain | Mass: 38951.105 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #38: RNA chain | Mass: 50682.922 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #81: RNA chain | Mass: 6494.952 Da / Num. of mol.: 1 / Source method: obtained synthetically / Details: synthetic mRNA / Source: (synth.) synthetic construct (others) |
| #82: RNA chain | Mass: 24890.121 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Details: N-acetylated Phe-tRNA / Source: (natural) |
+60S ribosomal protein ... , 40 types, 40 molecules a5A5b5B5c5C5D5d5e5f5F5g5G5h5H5i5I5J5j5k5l5L5M5N5o5p5P5Q5S5U5...
-Non-polymers , 5 types, 1081 molecules 

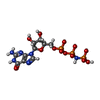

| #83: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 8 / Source method: obtained synthetically / Formula: Zn#84: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 1058 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 1058 / Source method: obtained synthetically / Formula: Mg#85: Chemical | ChemComp-GNP / |  Mass: 522.196 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H17N6O13P3 Mass: 522.196 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H17N6O13P3Comment: GppNHp, GMPPNP, energy-carrying molecule analogue*YM #86: Chemical | ChemComp-5CR / |  Type: L-peptide linking / Mass: 207.226 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C11H13NO3 Type: L-peptide linking / Mass: 207.226 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C11H13NO3#87: Water | ChemComp-HOH / | Mass: 18.015 Da / Num. of mol.: 13 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Nonstop ribosomal complex bound with Dom34-Hbs1-GMPPNP Type: RIBOSOME / Entity ID: #1-#82 / Source: MULTIPLE SOURCES |
|---|---|
| Molecular weight | Value: 3.6 MDa / Experimental value: NO |
| Buffer solution | pH: 7.6 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Tecnai Polara / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI POLARA 300 |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 80645 X / Nominal defocus max: 4500 nm / Nominal defocus min: 500 nm / Cs: 2 mm |
| Specimen holder | Cryogen: NITROGEN |
| Image recording | Average exposure time: 5 sec. / Electron dose: 25 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 SUMMIT (4k x 4k) / Num. of real images: 4797 |
| Image scans | Movie frames/image: 25 / Used frames/image: 1-25 |
- Processing
Processing
| Software | Name: PHENIX / Version: dev_2415: / Classification: refinement | ||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 520612 | ||||||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.3 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 73391 / Algorithm: FOURIER SPACE / Num. of class averages: 1 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL | ||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Highest resolution: 3.3 Å | ||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|
