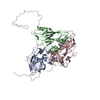
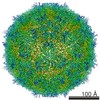
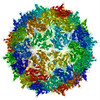
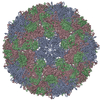
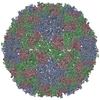
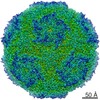
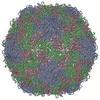
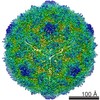
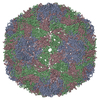
ジャーナル: J Virol / 年: 2017 タイトル: Cryo-electron Microscopy Study of the Genome Release of the Dicistrovirus Israeli Acute Bee Paralysis Virus. 著者: Edukondalu Mullapudi / Tibor Füzik / Antonín Přidal / Pavel Plevka / 要旨: Viruses of the family Dicistroviridae can cause substantial economic damage by infecting agriculturally important insects. Israeli acute bee paralysis virus (IAPV) causes honeybee colony collapse ...Viruses of the family Dicistroviridae can cause substantial economic damage by infecting agriculturally important insects. Israeli acute bee paralysis virus (IAPV) causes honeybee colony collapse disorder in the United States. High-resolution molecular details of the genome delivery mechanism of dicistroviruses are unknown. Here we present a cryo-electron microscopy analysis of IAPV virions induced to release their genomes in vitro We determined structures of full IAPV virions primed to release their genomes to a resolution of 3.3 Å and of empty capsids to a resolution of 3.9 Å. We show that IAPV does not form expanded A particles before genome release as in the case of related enteroviruses of the family Picornaviridae The structural changes observed in the empty IAPV particles include detachment of the VP4 minor capsid proteins from the inner face of the capsid and partial loss of the structure of the N-terminal arms of the VP2 capsid proteins. Unlike the case for many picornaviruses, the empty particles of IAPV are not expanded relative to the native virions and do not contain pores in their capsids that might serve as channels for genome release. Therefore, rearrangement of a unique region of the capsid is probably required for IAPV genome release. IMPORTANCE: Honeybee populations in Europe and North America are declining due to pressure from pathogens, including viruses. Israeli acute bee paralysis virus (IAPV), a member of the family ...IMPORTANCE: Honeybee populations in Europe and North America are declining due to pressure from pathogens, including viruses. Israeli acute bee paralysis virus (IAPV), a member of the family Dicistroviridae, causes honeybee colony collapse disorder in the United States. The delivery of virus genomes into host cells is necessary for the initiation of infection. Here we present a structural cryo-electron microscopy analysis of IAPV particles induced to release their genomes. We show that genome release is not preceded by an expansion of IAPV virions as in the case of related picornaviruses that infect vertebrates. Furthermore, minor capsid proteins detach from the capsid upon genome release. The genome leaves behind empty particles that have compact protein shells.
履歴
登録
2016年9月16日
登録サイト: PDBE / 処理サイト: PDBE
改定 1.0
2016年11月30日
Provider: repository / タイプ: Initial release
改定 1.1
2016年12月21日
Group: Database references
改定 1.2
2017年2月8日
Group: Database references
改定 1.3
2017年8月2日
Group: Data collection / カテゴリ: em_software / Item: _em_software.name
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Israeli acute paralysis virus (ウイルス)
Israeli acute paralysis virus (ウイルス) データ登録者
データ登録者 チェコ,
チェコ,  ドイツ, 2件
ドイツ, 2件  引用
引用 構造の表示
構造の表示 UCSF Chimera
UCSF Chimera ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード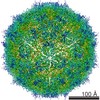




 PDBj
PDBj
 集合体
集合体