 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
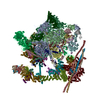
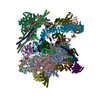
Yorodumi- PDB-5lj5: Overall structure of the yeast spliceosome immediately after bran... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5lj5 | ||||||
|---|---|---|---|---|---|---|---|
| Title | Overall structure of the yeast spliceosome immediately after branching. | ||||||
 Components Components |
| ||||||
 Keywords Keywords | SPLICING / spliceosome / snRNP / pre-mRNA splicing / trans-esterification / lariat intermediate / complex C | ||||||
| Function / homology |  Function and homology information Function and homology informationRNA exon ligation / first spliceosomal transesterification activity / snRNA metabolic process / snRNA modification / post-spliceosomal complex / U2-type post-mRNA release spliceosomal complex / cellular bud site selection / post-mRNA release spliceosomal complex / generation of catalytic spliceosome for first transesterification step / 3'-5' RNA helicase activity ...RNA exon ligation / first spliceosomal transesterification activity / snRNA metabolic process / snRNA modification / post-spliceosomal complex / U2-type post-mRNA release spliceosomal complex / cellular bud site selection / post-mRNA release spliceosomal complex / generation of catalytic spliceosome for first transesterification step / 3'-5' RNA helicase activity / cis assembly of pre-catalytic spliceosome / 7-methylguanosine cap hypermethylation / pICln-Sm protein complex / U2-type catalytic step 1 spliceosome / ATP-dependent activity, acting on RNA / small nuclear ribonucleoprotein complex / SMN-Sm protein complex / splicing factor binding / spliceosomal tri-snRNP complex / commitment complex / U4 snRNP / snRNP binding / mRNA cis splicing, via spliceosome / U2-type prespliceosome assembly / U2-type catalytic step 2 spliceosome / U2-type spliceosomal complex / pre-mRNA binding / spliceosome conformational change to release U4 (or U4atac) and U1 (or U11) / U1 snRNP / U2 snRNP / U2-type prespliceosome / poly(U) RNA binding / generation of catalytic spliceosome for second transesterification step / precatalytic spliceosome / mRNA 5'-splice site recognition / mRNA 3'-splice site recognition / spliceosomal complex assembly / Gap-filling DNA repair synthesis and ligation in TC-NER / Prp19 complex / DNA replication origin binding / spliceosomal tri-snRNP complex assembly / U5 snRNP / Dual incision in TC-NER / U5 snRNA binding / DNA replication initiation / pre-mRNA intronic binding / U2 snRNA binding / U6 snRNA binding / U1 snRNA binding / protein K63-linked ubiquitination / U4/U6 x U5 tri-snRNP complex / spliceosomal snRNP assembly / catalytic step 2 spliceosome / positive regulation of cell cycle / positive regulation of RNA splicing / spliceosomal complex / mRNA splicing, via spliceosome / RING-type E3 ubiquitin transferase / metallopeptidase activity / ubiquitin-protein transferase activity / ubiquitin protein ligase activity / RNA helicase activity / RNA helicase / DNA repair / mRNA binding / GTPase activity / chromatin binding / GTP binding / chromatin / ATP hydrolysis activity / mitochondrion / DNA binding / RNA binding / zinc ion binding / ATP binding / metal ion binding / identical protein binding / nucleus / cytosol / cytoplasm Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 10 Å | ||||||
 Authors Authors | Galej, W.P. / Wilkinson, M.F. / Fica, S.M. / Oubridge, C. / Newman, A.J. / Nagai, K. | ||||||
 Citation Citation | Journal: Nature / Year: 2016 Title: Cryo-EM structure of the spliceosome immediately after branching. Authors: Wojciech P Galej / Max E Wilkinson / Sebastian M Fica / Chris Oubridge / Andrew J Newman / Kiyoshi Nagai /  Abstract: Precursor mRNA (pre-mRNA) splicing proceeds by two consecutive transesterification reactions via a lariat-intron intermediate. Here we present the 3.8 Å cryo-electron microscopy structure of the ...Precursor mRNA (pre-mRNA) splicing proceeds by two consecutive transesterification reactions via a lariat-intron intermediate. Here we present the 3.8 Å cryo-electron microscopy structure of the spliceosome immediately after lariat formation. The 5'-splice site is cleaved but remains close to the catalytic Mg site in the U2/U6 small nuclear RNA (snRNA) triplex, and the 5'-phosphate of the intron nucleotide G(+1) is linked to the branch adenosine 2'OH. The 5'-exon is held between the Prp8 amino-terminal and linker domains, and base-pairs with U5 snRNA loop 1. Non-Watson-Crick interactions between the branch helix and 5'-splice site dock the branch adenosine into the active site, while intron nucleotides +3 to +6 base-pair with the U6 snRNA ACAGAGA sequence. Isy1 and the step-one factors Yju2 and Cwc25 stabilize docking of the branch helix. The intron downstream of the branch site emerges between the Prp8 reverse transcriptase and linker domains and extends towards the Prp16 helicase, suggesting a plausible mechanism of remodelling before exon ligation. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5lj5.cif.gz | 2.4 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5lj5.ent.gz | 1.7 MB | Display | PDB format |
| PDBx/mmJSON format | 5lj5.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/lj/5lj5ftp://data.pdbj.org/pub/pdb/validation_reports/lj/5lj5 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4057MC  4055C  4056C  4058C  4059C  5lj3C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data | |
| EM raw data | EMPIAR-10687 (Title: Yeast C, Ci, C*, and P complex spliceosomes / Data size: 8.9 TB Data #1: Unaligned movies of C-complex spliceosome with 3' splice site AG to AC mutation (Dataset 1) [micrographs - multiframe] Data #2: Unaligned movies of C and C*-complex spliceosomes with 3' splice site AG to AdG mutation (Dataset 2) [micrographs - multiframe] Data #3: Unaligned movies of C and C*-complex spliceosomes with 3' splice site AG to AdG mutation (Dataset 3) [micrographs - multiframe] Data #4: Aligned movies of C-complex spliceosomes with cold-sensitive prp16-302 mutation, purified with Cwc25 (Dataset 4) [micrographs - multiframe] Data #5: Unaligned movies of C-complex spliceosomes with cold-sensitive prp16-302 mutation, purified with Cwc25 and incubated with ATP and Mg (Dataset 5) [micrographs - multiframe] Data #6: Unaligned movies of C, C*, and P-complex spliceosomes with dominant-negative Prp22 mutation K512A, purified with Slu7 (Dataset 6) [micrographs - multiframe] Data #7: Unaligned movies of P-complex spliceosomes with dominant-negative Prp22 mutation K512A, treated with anti-3'exon RNaseH oligo, purified in presence of Mg (Dataset 9) [micrographs - single frame] Data #8: Selected C-complex particles after polishing [picked particles - single frame - processed] Data #9: Selected P-complex particles after polishing [picked particles - single frame - processed] Data #10: Various signal subtractions for C- and P-complex spliceosomes [picked particles - single frame - processed]) |







-Links
 PDBj
PDBj




































- Assembly
Assembly
| Deposited unit | 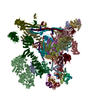
|
|---|---|
| 1 |
|
-Components
-RNA chain , 5 types, 5 molecules UEIZV
| #1: RNA chain | Mass: 57444.875 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #2: RNA chain | Mass: 5170.152 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #3: RNA chain | Mass: 24109.115 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #4: RNA chain | Mass: 376267.406 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #5: RNA chain | Mass: 35883.176 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Pre-mRNA-splicing factor ... , 14 types, 14 molecules AFCGJLMNORSTQs
| #6: Protein | Mass: 279867.469 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #9: Protein | Mass: 20412.477 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #10: Protein | Mass: 114174.008 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #11: Protein | Mass: 28073.836 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #13: Protein | Mass: 50771.289 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #15: Protein | Mass: 18484.502 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #16: Protein | Mass: 38486.562 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #17: Protein | Mass: 40988.590 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #18: Protein | Mass: 67837.773 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #20: Protein | Mass: 15793.596 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #21: Protein | Mass: 82555.859 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #22: Protein | Mass: 100344.016 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #32: Protein | Mass: 121815.234 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #34: Protein | Mass: 20741.455 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Protein , 6 types, 7 molecules BDHPbkx
| #7: Protein | Mass: 246470.266 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||
|---|---|---|---|
| #8: Protein | Mass: 32371.086 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||
| #12: Protein | Mass: 69118.984 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||
| #19: Protein | Mass: 19970.195 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||
| #23: Protein | Mass: 22426.990 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #35: Protein | | Mass: 16017.708 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Pre-mRNA-processing ... , 2 types, 5 molecules Ktuvw
| #14: Protein | Mass: 42548.727 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #33: Protein | Mass: 56629.777 Da / Num. of mol.: 4 / Source method: isolated from a natural source / Source: (natural) References: UniProt: P32523, Ligases; Forming carbon-nitrogen bonds; Acid-amino-acid ligases (peptide synthases) |
-Small nuclear ribonucleoprotein ... , 6 types, 12 molecules dnepfqgrhljm
| #24: Protein | Mass: 11240.139 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #25: Protein | Mass: 10385.098 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #26: Protein | Mass: 9669.945 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #27: Protein | Mass: 8490.809 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #28: Protein | Mass: 16296.798 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #29: Protein | Mass: 12876.066 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) |
|---|
-U2 small nuclear ribonucleoprotein ... , 2 types, 2 molecules WY
| #30: Protein | Mass: 27232.252 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #31: Protein | Mass: 12850.944 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-Non-polymers , 3 types, 10 molecules 

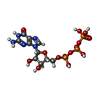
| #36: Chemical |  Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg#37: Chemical | ChemComp-ZN /  Mass: 65.409 Da / Num. of mol.: 7 / Source method: obtained synthetically / Formula: Zn Mass: 65.409 Da / Num. of mol.: 7 / Source method: obtained synthetically / Formula: Zn#38: Chemical | ChemComp-GTP / |  Mass: 523.180 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM Mass: 523.180 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM |
|---|
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Spliceosome immediately after branching / Type: COMPLEX Details: Splicing extract was prepared from Prp18-HA or Slu7-TAPS yeast strains. An in vitro transcribed yeast UBC4 pre-mRNA substrate (with 2 x MS2 bacteriophage coat protein-binding stem loops at ...Details: Splicing extract was prepared from Prp18-HA or Slu7-TAPS yeast strains. An in vitro transcribed yeast UBC4 pre-mRNA substrate (with 2 x MS2 bacteriophage coat protein-binding stem loops at the 5' end and with the 3'-splice site sequence UAGAG mutated to UACAC) was pre-bound to an MS2-maltose binding protein fusion protein. This substrate-protein complex was added to the splicing extract. The splicing reaction proceeded through the first step but the second step was blocked by the 3' splice site mutation. Substrate-bound spliceosomes from the splicing extract were purified on amylose resin and eluted with maltose. Subsequently the spliceosomes were captured on anti-HA-agarose (for Prp18-HA-tagged) or streptactin resin (for Slu7-TAPS tagged) and eluted with HA peptide or desthiobiotin, respectively. Purified spliceosomes were then dialysed against 20 mM HEPES KOH pH 7.8, 75 mM KCl, 0.25 mM EDTA Entity ID: #1-#35 / Source: NATURAL | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 2 MDa / Experimental value: NO | ||||||||||||||||||||||||||||||
| Source (natural) | Organism: | ||||||||||||||||||||||||||||||
| Buffer solution | pH: 7.8 | ||||||||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||||||||
| Specimen | Conc.: 0.3 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||
| Specimen support | Grid material: COPPER / Grid mesh size: 400 divisions/in. / Grid type: Quantifoil R2/2 | ||||||||||||||||||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK III / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K Details: 3 microlitres sample were applied to the grid, left for 30 seconds and then blotted for 2.5-3.0 seconds before plunging. |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 81000 X / Calibrated magnification: 35714 X / Nominal defocus max: 4000 nm / Nominal defocus min: 500 nm |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 0.8 sec. / Electron dose: 2 e/Å2 / Detector mode: SUPER-RESOLUTION / Film or detector model: GATAN K2 QUANTUM (4k x 4k) / Num. of real images: 2213 Details: Total dose: 40 electrons/Angstrom^2 over 16 seconds. 20 movie frames collected at 1.25 frames per second. |
| EM imaging optics | Energyfilter name: GIF Quantum |
| Image scans | Movie frames/image: 20 / Used frames/image: 1-20 |
- Processing
Processing
| Software | Name: REFMAC / Version: 5.8.0124 / Classification: refinement | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 15872 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 10 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 15872 / Num. of class averages: 1 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: OTHER / Space: RECIPROCAL Details: Used secondary structure restraints generated in ProSMART and LibG. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement | Resolution: 10→10 Å / Cor.coef. Fo:Fc: 0.954 / SU B: 27.594 / SU ML: 0.398 / ESU R: 0.522 Stereochemistry target values: MAXIMUM LIKELIHOOD WITH PHASES Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Solvent model: PARAMETERS FOR MASK CACLULATION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 180.1 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: 1 / Total: 63166 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
|