 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5h5u | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Mechanistic insights into the alternative translation termination by ArfA and RF2 | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | RIBOSOME / translation / RF2 / Arfa / non-stop mRNA / ribosome rescue | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報translation release factor activity, codon specific / stringent response / ribosomal large subunit binding / transcription antitermination factor activity, RNA binding / ornithine decarboxylase inhibitor activity / misfolded RNA binding / Group I intron splicing / RNA folding / transcriptional attenuation / endoribonuclease inhibitor activity ...translation release factor activity, codon specific / stringent response / ribosomal large subunit binding / transcription antitermination factor activity, RNA binding / ornithine decarboxylase inhibitor activity / misfolded RNA binding / Group I intron splicing / RNA folding / transcriptional attenuation / endoribonuclease inhibitor activity / positive regulation of ribosome biogenesis / RNA-binding transcription regulator activity / translational termination / four-way junction DNA binding / negative regulation of cytoplasmic translation / DnaA-L2 complex / regulation of mRNA stability / translation repressor activity / negative regulation of translational initiation / negative regulation of DNA-templated DNA replication initiation / rescue of stalled cytosolic ribosome / mRNA regulatory element binding translation repressor activity / positive regulation of RNA splicing / regulation of DNA-templated transcription elongation / transcription elongation factor complex / cytosolic ribosome assembly / response to reactive oxygen species / ribosome assembly / assembly of large subunit precursor of preribosome / transcription antitermination / DNA endonuclease activity / regulation of cell growth / DNA-templated transcription termination / response to radiation / maintenance of translational fidelity / mRNA 5'-UTR binding / regulation of translation / large ribosomal subunit / ribosomal small subunit assembly / transferase activity / ribosome biogenesis / ribosome binding / ribosomal small subunit biogenesis / 5S rRNA binding / small ribosomal subunit / ribosomal large subunit assembly / small ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / large ribosomal subunit rRNA binding / cytosolic large ribosomal subunit / cytoplasmic translation / tRNA binding / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / viral translational frameshifting / response to antibiotic / negative regulation of DNA-templated transcription / hydrolase activity / mRNA binding / DNA binding / RNA binding / zinc ion binding / membrane / cytoplasm / cytosol 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.01 Å | ||||||
 データ登録者 データ登録者 | Ma, C. / Kurita, D. / Li, N. / Chen, Y. / Himeno, H. / Gao, N. | ||||||
 引用 引用 | ジャーナル: Nature / 年: 2017 タイトル: Mechanistic insights into the alternative translation termination by ArfA and RF2. 著者: Chengying Ma / Daisuke Kurita / Ningning Li / Yan Chen / Hyouta Himeno / Ning Gao /   要旨: During cellular translation of messenger RNAs by ribosomes, the translation apparatus sometimes pauses or stalls at the elongation and termination steps. With the exception of programmed stalling, ...During cellular translation of messenger RNAs by ribosomes, the translation apparatus sometimes pauses or stalls at the elongation and termination steps. With the exception of programmed stalling, which is usually used by cells for regulatory purposes, ribosomes stalled on mRNAs need to be terminated and recycled to maintain adequate translation capacity. Much ribosome stalling originates in aberrant mRNAs that lack a stop codon. Transcriptional errors, misprocessing of primary transcripts, and undesired mRNA cleavage all contribute to the formation of non-stop mRNAs. Ribosomes stalled at the 3' end of non-stop mRNAs do not undergo normal termination owing to the lack of specific stop-codon recognition by canonical peptide release factors at the A-site decoding centre. In bacteria, the transfer-messenger RNA (tmRNA)-SmpB-mediated trans-translation rescue system reroutes stalled ribosomes to the normal elongation cycle and translation termination. Two additional rescue systems, ArfA-RF2 (refs 13, 14, 15, 16) and ArfB (formerly known as YaeJ), are also present in many bacterial species, but their mechanisms are not fully understood. Here, using cryo-electron microscopy, we characterize the structure of the Escherichia coli 70S ribosome bound with ArfA, the release factor RF2, a short non-stop mRNA and a cognate P-site tRNA. The C-terminal loop of ArfA occupies the mRNA entry channel on the 30S subunit, whereas its N terminus is sandwiched between the decoding centre and the switch loop of RF2, leading to marked conformational changes in both the decoding centre and RF2. Despite the distinct conformation of RF2, its conserved catalytic GGQ motif is precisely positioned next to the CCA-end of the P-site tRNA. These data illustrate a stop-codon surrogate mechanism for ArfA in facilitating the termination of non-stop ribosomal complexes by RF2. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5h5u.cif.gz | 3.7 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5h5u.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 5h5u.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/h5/5h5uftp://data.pdbj.org/pub/pdb/validation_reports/h5/5h5u | HTTPS FTP |
|---|
-関連構造データ




-リンク
 PDBj
PDBj





























- 集合体
集合体
| 登録構造単位 | 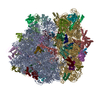
|
|---|---|
| 1 |
|
-要素
-30S ribosomal protein ... , 20種, 20分子 12ijklmnopqrstuvwxyz
| #1: タンパク質 | 分子量: 9577.268 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: タンパク質 | 分子量: 8392.844 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #40: タンパク質 | 分子量: 26650.475 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #41: タンパク質 | 分子量: 25900.117 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #42: タンパク質 | 分子量: 23383.002 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #43: タンパク質 | 分子量: 17498.203 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #44: タンパク質 | 分子量: 15727.512 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #45: タンパク質 | 分子量: 19923.959 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #46: タンパク質 | 分子量: 14015.361 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #47: タンパク質 | 分子量: 14755.074 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #48: タンパク質 | 分子量: 11755.597 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #49: タンパク質 | 分子量: 13739.778 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #50: タンパク質 | 分子量: 13636.961 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #51: タンパク質 | 分子量: 12997.271 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #52: タンパク質 | 分子量: 11475.364 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #53: タンパク質 | 分子量: 10159.621 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #54: タンパク質 | 分子量: 9207.572 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #55: タンパク質 | 分子量: 9593.296 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #56: タンパク質 | 分子量: 8874.276 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #57: タンパク質 | 分子量: 10324.160 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 2種, 2分子 34
| #3: タンパク質 | 分子量: 8190.438 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #4: タンパク質 | 分子量: 41300.660 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-RNA鎖 , 5種, 5分子 57ABh
| #5: RNA鎖 | 分子量: 24497.602 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) |
|---|---|
| #6: RNA鎖 | 分子量: 1876.173 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) |
| #7: RNA鎖 | 分子量: 941305.188 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #8: RNA鎖 | 分子量: 38790.090 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #39: RNA鎖 | 分子量: 497221.594 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+50S ribosomal protein ... , 30種, 30分子 CDEFGHIJKLMNOPQRSTUVWXYZabcdef
-詳細
| Has protein modification | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 由来(天然) |
| ||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||
| 緩衝液 | pH: 7.6 | ||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER | ||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277.15 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 電子線照射量: 2 e/Å2 / 検出モード: INTEGRATING フィルム・検出器のモデル: FEI FALCON II (4k x 4k) |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0088 / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: NONE | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.01 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 13693 / クラス平均像の数: 8 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 3.01→3.01 Å / Cor.coef. Fo:Fc: 0.971 / SU B: 9.616 / SU ML: 0.159 / ESU R: 0.249 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | イオンプローブ半径: 0.8 Å / 減衰半径: 0.8 Å / VDWプローブ半径: 1.2 Å / 溶媒モデル: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 144.877 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 合計: 149802 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|