 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-5flz: Cryo-EM structure of gamma-TuSC oligomers in a closed conformation -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 5flz | ||||||
|---|---|---|---|---|---|---|---|
| Title | Cryo-EM structure of gamma-TuSC oligomers in a closed conformation | ||||||
 Components Components |
| ||||||
 Keywords Keywords | CELL CYCLE / MICROTUBULE NUCLEATION | ||||||
| Function / homology |  Function and homology information Function and homology informationinner plaque of spindle pole body / microtubule nucleation by spindle pole body / outer plaque of spindle pole body / gamma-tubulin small complex / regulation of microtubule nucleation / microtubule nucleator activity / mitotic spindle pole body / gamma-tubulin complex / gamma-tubulin ring complex / meiotic spindle organization ...inner plaque of spindle pole body / microtubule nucleation by spindle pole body / outer plaque of spindle pole body / gamma-tubulin small complex / regulation of microtubule nucleation / microtubule nucleator activity / mitotic spindle pole body / gamma-tubulin complex / gamma-tubulin ring complex / meiotic spindle organization / microtubule nucleation / spindle pole body / gamma-tubulin binding / positive regulation of cytoplasmic translation / mitotic sister chromatid segregation / spindle assembly / cytoplasmic microtubule organization / mitotic spindle organization / meiotic cell cycle / structural constituent of cytoskeleton / spindle / spindle pole / mitotic cell cycle / microtubule / GTP binding / nucleus / cytoplasm Similarity search - Function | ||||||
| Biological species |  | ||||||
| Method | ELECTRON MICROSCOPY / helical reconstruction / cryo EM / Resolution: 6.9 Å | ||||||
 Authors Authors | Greenberg, C.H. / Kollman, J. / Zelter, A. / Johnson, R. / MacCoss, M.J. / Davis, T.N. / Agard, D.A. / Sali, A. | ||||||
 Citation Citation | Journal: J Struct Biol / Year: 2016 Title: Structure of γ-tubulin small complex based on a cryo-EM map, chemical cross-links, and a remotely related structure. Authors: Charles H Greenberg / Justin Kollman / Alex Zelter / Richard Johnson / Michael J MacCoss / Trisha N Davis / David A Agard / Andrej Sali /  Abstract: Modeling protein complex structures based on distantly related homologues can be challenging due to poor sequence and structure conservation. Therefore, utilizing even low-resolution experimental ...Modeling protein complex structures based on distantly related homologues can be challenging due to poor sequence and structure conservation. Therefore, utilizing even low-resolution experimental data can significantly increase model precision and accuracy. Here, we present models of the two key functional states of the yeast γ-tubulin small complex (γTuSC): one for the low-activity "open" state and another for the higher-activity "closed" state. Both models were computed based on remotely related template structures and cryo-EM density maps at 6.9Å and 8.0Å resolution, respectively. For each state, extensive sampling of alignments and conformations was guided by the fit to the corresponding cryo-EM density map. The resulting good-scoring models formed a tightly clustered ensemble of conformations in most regions. We found significant structural differences between the two states, primarily in the γ-tubulin subunit regions where the microtubule binds. We also report a set of chemical cross-links that were found to be consistent with equilibrium between the open and closed states. The protocols developed here have been incorporated into our open-source Integrative Modeling Platform (IMP) software package (http://integrativemodeling.org), and can therefore be applied to many other systems. #1: Journal: Nat Struct Mol Biol / Year: 2015Title: Ring closure activates yeast γTuRC for species-specific microtubule nucleation. Authors: Justin M Kollman / Charles H Greenberg / Sam Li / Michelle Moritz / Alex Zelter / Kimberly K Fong / Jose-Jesus Fernandez / Andrej Sali / John Kilmartin / Trisha N Davis / David A Agard /   Abstract: The γ-tubulin ring complex (γTuRC) is the primary microtubule nucleator in cells. γTuRC is assembled from repeating γ-tubulin small complex (γTuSC) subunits and is thought to function as a ...The γ-tubulin ring complex (γTuRC) is the primary microtubule nucleator in cells. γTuRC is assembled from repeating γ-tubulin small complex (γTuSC) subunits and is thought to function as a template by presenting a γ-tubulin ring that mimics microtubule geometry. However, a previous yeast γTuRC structure showed γTuSC in an open conformation that prevents matching to microtubule symmetry. By contrast, we show here that γ-tubulin complexes are in a closed conformation when attached to microtubules. To confirm the functional importance of the closed γTuSC ring, we trapped the closed state and determined its structure, showing that the γ-tubulin ring precisely matches microtubule symmetry and providing detailed insight into γTuRC architecture. Importantly, the closed state is a stronger nucleator, thus suggesting that this conformational switch may allosterically control γTuRC activity. Finally, we demonstrate that γTuRCs have a strong preference for tubulin from the same species. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 5flz.cif.gz | 3.4 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb5flz.ent.gz | 2.8 MB | Display | PDB format |
| PDBx/mmJSON format | 5flz.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 5flz_validation.pdf.gz | 1.3 MB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 5flz_full_validation.pdf.gz | 2.2 MB | Display | |
| Data in XML | 5flz_validation.xml.gz | 487.2 KB | Display | |
| Data in CIF | 5flz_validation.cif.gz | 720.3 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/fl/5flzftp://data.pdbj.org/pub/pdb/validation_reports/fl/5flz | HTTPS FTP |
-Related structure data
| Related structure data |  2799M  5fm1C C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |


-Links
 PDBj
PDBj


- Assembly
Assembly
| Deposited unit | 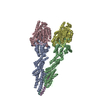
|
|---|---|
| 1 | x 7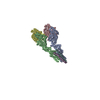
|
| Number of models | 10 |
-Components
| #1: Protein | Mass: 96940.594 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Production host:  UNIDENTIFIED BACULOVIRUS / References: UniProt: P38863 UNIDENTIFIED BACULOVIRUS / References: UniProt: P38863 | ||||
|---|---|---|---|---|---|
| #2: Protein | Mass: 98336.211 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Production host: UNIDENTIFIED BACULOVIRUS / References: UniProt: P53540 | ||||
| #3: Protein | Mass: 52733.340 Da / Num. of mol.: 2 / Mutation: YES Source method: isolated from a genetically manipulated source Details: CYSTEINE RESIDUES WERE INTRODUCED AT POSITIONS 58 AND 288 TO PROMOTE CROSSLINKING OF THE HELICAL COMPLEX Source: (gene. exp.) Production host: UNIDENTIFIED BACULOVIRUS / References: UniProt: P53378#4: Protein/peptide | Mass: 3762.629 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Details: RESIDUES 1-220 OF SPC110 WERE EXPRESSED WITH AN N-TERMINAL GST TAG, WHICH WAS CLEAVED OFF DURING PURIFICATION. IT CANNOT BE UNIQUELY IDENTIFIED WHICH FRAGMENT OF SPC110 IS PRESENT IN THE ...Details: RESIDUES 1-220 OF SPC110 WERE EXPRESSED WITH AN N-TERMINAL GST TAG, WHICH WAS CLEAVED OFF DURING PURIFICATION. IT CANNOT BE UNIQUELY IDENTIFIED WHICH FRAGMENT OF SPC110 IS PRESENT IN THE MAP, THUS A POLY-ALANINE HELIX WAS FITTED INTO THE MAP. Source: (gene. exp.) Production host: UNIDENTIFIED BACULOVIRUSSequence details | CONTAINS TWO MUTATIONS TO FORCE CLOSURE, S58C AND G288C CHAINS E AND F CORRESPOND | |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: FILAMENT / 3D reconstruction method: helical reconstruction |
- Sample preparation
Sample preparation
| Component | Name: RECOMBINANT YEAST GAMMA- TUSC MUTANT S58C G288C / Type: COMPLEX |
|---|---|
| Buffer solution | Name: 40 MM HEPES PH 7.6, 100 MM KCL, 1 MM EGTA, 1MM MGCL2, 1 MM OXIDIZED GLUTATHIONE pH: 7.6 Details: 40 MM HEPES PH 7.6, 100 MM KCL, 1 MM EGTA, 1MM MGCL2, 1 MM OXIDIZED GLUTATHIONE |
| Specimen | Conc.: 2 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Details: OTHER |
| Vitrification | Instrument: FEI VITROBOT MARK I / Cryogen name: ETHANE Details: CRYOGEN- ETHANE, HUMIDITY- 90, INSTRUMENT- FEI VITROBOT MARK I, METHOD- BLOT FOR 2-5 SECONDS BEFORE PLUNGING, |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Tecnai F20 / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TECNAI F20 / Date: May 25, 2011 |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Calibrated magnification: 94000 X / Nominal defocus max: 2000 nm / Nominal defocus min: 800 nm / Cs: 2.12 mm |
| Image recording | Electron dose: 20 e/Å2 / Film or detector model: TVIPS TEMCAM-F816 (8k x 8k) |
| Image scans | Num. digital images: 364 |
- Processing
Processing
| EM software |
| |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Details: EACH MICROGRAPH | |||||||||||||||||||||||||||||||||||
| 3D reconstruction | Method: REFERENCE MATCHING / Resolution: 6.9 Å Details: INITIAL MODELS WERE GENERATED WITH MODELER 9.15 BASED ON PDB ENTRIES 3RIP (FOR GCP2 AND GCP3) AND 3CB2 (FOR GAMMA TUBULIN). TWO COMPLETE GAMMA-TUSC STRUCTURES WERE MODELED SIMULTANEOUSLY, ...Details: INITIAL MODELS WERE GENERATED WITH MODELER 9.15 BASED ON PDB ENTRIES 3RIP (FOR GCP2 AND GCP3) AND 3CB2 (FOR GAMMA TUBULIN). TWO COMPLETE GAMMA-TUSC STRUCTURES WERE MODELED SIMULTANEOUSLY, RIGIDLY FITTED INTO THE CRYO-EM MAP WITH UCSF CHIMERA, THEN FLEXIBLY FITTED WITH MDFF. ADDITIONAL SECONDARY STRUCTURE AND SYMMETRY RESTRAINTS WERE ADDED. FINAL VALIDATION PERFORMED WITH THE INTEGRATIVE MODELING PLATFORM. FOR DETAILS, ALL CODE IS DEPOSITED AT GITHUB.COM/INTEGRATIVEMODELING/GAMMA-TUSC SUBMISSION BASED ON EXPERIMENTAL DATA FROM EMDB EMD-2799. (DEPOSITION ID: 12869). Symmetry type: HELICAL | |||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT / Space: REAL / Details: METHOD--FLEXIBLE FITTING | |||||||||||||||||||||||||||||||||||
| Refinement | Highest resolution: 6.9 Å | |||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Highest resolution: 6.9 Å
|
