 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 4v8m | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
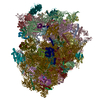
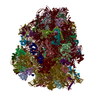
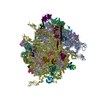
| タイトル | High-resolution cryo-electron microscopy structure of the Trypanosoma brucei ribosome | |||||||||||||||
 要素 要素 |
| |||||||||||||||
 キーワード キーワード | RIBOSOME / EUKARYOTIC / KINETOPLASTIDS / EXPANSION SEGMENTS | |||||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報organellar small ribosomal subunit / organellar large ribosomal subunit / ciliary transition zone / mitochondrial large ribosomal subunit / nuclear lumen / phosphate ion binding / negative regulation of translational frameshifting / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / protein-RNA complex assembly / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) ...organellar small ribosomal subunit / organellar large ribosomal subunit / ciliary transition zone / mitochondrial large ribosomal subunit / nuclear lumen / phosphate ion binding / negative regulation of translational frameshifting / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / protein-RNA complex assembly / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of LSU-rRNA / translation regulator activity / rescue of stalled ribosome / protein kinase C binding / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / ribosomal large subunit biogenesis / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / ribosome assembly / maturation of SSU-rRNA / small-subunit processome / regulation of cell growth / maintenance of translational fidelity / modification-dependent protein catabolic process / protein tag activity / rRNA processing / ribosome biogenesis / regulation of cell population proliferation / ribosome binding / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / 5S rRNA binding / small ribosomal subunit rRNA binding / ribosomal large subunit assembly / cytosolic small ribosomal subunit / large ribosomal subunit rRNA binding / cytosolic large ribosomal subunit / cytoplasmic translation / negative regulation of translation / rRNA binding / protein ubiquitination / structural constituent of ribosome / ribosome / translation / ribonucleoprotein complex / mRNA binding / ubiquitin protein ligase binding / nucleolus / RNA binding / zinc ion binding / nucleoplasm / nucleus / cytoplasm / cytosol 類似検索 - 分子機能 | |||||||||||||||
| 生物種 |  | |||||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 5.57 Å | |||||||||||||||
 データ登録者 データ登録者 | Hashem, Y. / des Georges, A. / Fu, J. / Buss, S.N. / Jossinet, F. / Jobe, A. / Zhang, Q. / Liao, H.Y. / Grassucci, R.A. / Bajaj, C. ...Hashem, Y. / des Georges, A. / Fu, J. / Buss, S.N. / Jossinet, F. / Jobe, A. / Zhang, Q. / Liao, H.Y. / Grassucci, R.A. / Bajaj, C. / Westhof, E. / Madison-Antenucci, S. / Frank, J. | |||||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2013 タイトル: High-resolution cryo-electron microscopy structure of the Trypanosoma brucei ribosome. 著者: Yaser Hashem / Amedee des Georges / Jie Fu / Sarah N Buss / Fabrice Jossinet / Amy Jobe / Qin Zhang / Hstau Y Liao / Robert A Grassucci / Chandrajit Bajaj / Eric Westhof / Susan Madison- ...著者: Yaser Hashem / Amedee des Georges / Jie Fu / Sarah N Buss / Fabrice Jossinet / Amy Jobe / Qin Zhang / Hstau Y Liao / Robert A Grassucci / Chandrajit Bajaj / Eric Westhof / Susan Madison-Antenucci / Joachim Frank /  要旨: Ribosomes, the protein factories of living cells, translate genetic information carried by messenger RNAs into proteins, and are thus involved in virtually all aspects of cellular development and ...Ribosomes, the protein factories of living cells, translate genetic information carried by messenger RNAs into proteins, and are thus involved in virtually all aspects of cellular development and maintenance. The few available structures of the eukaryotic ribosome reveal that it is more complex than its prokaryotic counterpart, owing mainly to the presence of eukaryote-specific ribosomal proteins and additional ribosomal RNA insertions, called expansion segments. The structures also differ among species, partly in the size and arrangement of these expansion segments. Such differences are extreme in kinetoplastids, unicellular eukaryotic parasites often infectious to humans. Here we present a high-resolution cryo-electron microscopy structure of the ribosome of Trypanosoma brucei, the parasite that is transmitted by the tsetse fly and that causes African sleeping sickness. The atomic model reveals the unique features of this ribosome, characterized mainly by the presence of unusually large expansion segments and ribosomal-protein extensions leading to the formation of four additional inter-subunit bridges. We also find additional rRNA insertions, including one large rRNA domain that is not found in other eukaryotes. Furthermore, the structure reveals the five cleavage sites of the kinetoplastid large ribosomal subunit (LSU) rRNA chain, which is known to be cleaved uniquely into six pieces, and suggests that the cleavage is important for the maintenance of the T. brucei ribosome in the observed structure. We discuss several possible implications of the large rRNA expansion segments for the translation-regulation process. The structure could serve as a basis for future experiments aimed at understanding the functional importance of these kinetoplastid-specific ribosomal features in protein-translation regulation, an essential step towards finding effective and safe kinetoplastid-specific drugs. | |||||||||||||||
| 履歴 |
| |||||||||||||||
| Remark 700 | SHEET DETERMINATION METHOD: DSSP THE SHEETS PRESENTED AS "5B" IN EACH CHAIN ON SHEET RECORDS BELOW ... SHEET DETERMINATION METHOD: DSSP THE SHEETS PRESENTED AS "5B" IN EACH CHAIN ON SHEET RECORDS BELOW IS ACTUALLY AN 5-STRANDED BARREL THIS IS REPRESENTED BY A 6-STRANDED SHEET IN WHICH THE FIRST AND LAST STRANDS ARE IDENTICAL. THE SHEETS PRESENTED AS "EA" IN EACH CHAIN ON SHEET RECORDS BELOW IS ACTUALLY AN 5-STRANDED BARREL THIS IS REPRESENTED BY A 6-STRANDED SHEET IN WHICH THE FIRST AND LAST STRANDS ARE IDENTICAL. THE SHEETS PRESENTED AS "SA" IN EACH CHAIN ON SHEET RECORDS BELOW IS ACTUALLY AN 5-STRANDED BARREL THIS IS REPRESENTED BY A 6-STRANDED SHEET IN WHICH THE FIRST AND LAST STRANDS ARE IDENTICAL. |
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 4v8m.cif.gz | 5.6 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb4v8m.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 4v8m.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 4v8m_validation.pdf.gz | 1.9 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 4v8m_full_validation.pdf.gz | 7.8 MB | 表示 | |
| XML形式データ | 4v8m_validation.xml.gz | 929.7 KB | 表示 | |
| CIF形式データ | 4v8m_validation.cif.gz | 1.3 MB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/v8/4v8mftp://data.pdbj.org/pub/pdb/validation_reports/v8/4v8m | HTTPS FTP |
-関連構造データ

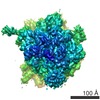



-リンク
 PDBj
PDBj






























- 集合体
集合体
| 登録構造単位 | 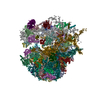
|
|---|---|
| 1 |
|
-要素
+40S RIBOSOMAL PROTEIN ... , 25種, 25分子 A0A1A2A3A5A6ACADAFAGAHAIAJAKALAMAPARASATAUAWAXAYAZ
-RIBOSOMAL PROTEIN ... , 9種, 9分子 A4A8AOAQAVBQBfBmBn
| #5: タンパク質 | 分子量: 23889.201 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q38FD9 |
|---|---|
| #9: タンパク質 | 分子量: 6679.840 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 |
| #22: タンパク質 | 分子量: 18891.506 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q583K7 |
| #24: タンパク質 | 分子量: 13248.478 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 |
| #29: タンパク質 | 分子量: 12837.207 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q384J9 |
| #50: タンパク質 | 分子量: 26461.006 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q38CY7 |
| #65: タンパク質 | 分子量: 48802.996 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q581Q1 |
| #72: タンパク質 | 分子量: 12491.966 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q38CY6 |
| #73: タンパク質 | 分子量: 10128.931 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q386L1 |
-タンパク質 , 7種, 7分子 A7A9AEBJBPBhBs
| #8: タンパク質 | 分子量: 34727.836 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q383E1 |
|---|---|
| #10: タンパク質 | 分子量: 16936.711 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q57WI3 |
| #13: タンパク質 | 分子量: 20132.459 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q38CI4 |
| #43: タンパク質 | 分子量: 24640.195 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q383V8 |
| #49: タンパク質 | 分子量: 21742.795 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: P55842 |
| #67: タンパク質 | 分子量: 21752.922 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: Q38DH8 |
| #78: タンパク質 | 分子量: 14685.345 Da / 分子数: 1 / 由来タイプ: 天然 由来: (天然) 株: 927/4 GUTAT10.1 / 参照: UniProt: P21899 |
-RNA鎖 , 10種, 10分子 BABBBCBDBEBFBGBHAAAB
| #34: RNA鎖 | 分子量: 595796.438 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #35: RNA鎖 | 分子量: 471450.062 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #36: RNA鎖 | 分子量: 54224.051 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #37: RNA鎖 | 分子量: 38316.672 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #38: RNA鎖 | 分子量: 67234.633 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #39: RNA鎖 | 分子量: 23079.566 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #40: RNA鎖 | 分子量: 58889.910 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #41: RNA鎖 | 分子量: 43388.613 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #85: RNA鎖 | 分子量: 724377.062 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #86: RNA鎖 | 分子量: 23545.971 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+60S RIBOSOMAL PROTEIN ... , 35種, 35分子 BIBKBLBMBNBOBRBSBTBUBVBWBXBYBZBaBbBcBdBeBgBiBjBkBlBoBpBqBrBt...
-詳細
| Has protein modification | Y |
|---|---|
| 配列の詳細 | CHAINS 8 AND Q ARE MODELLED USING T. CRUZI SEQUENCE UNP ACCESSIONS |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: TRYPANOSOMA BRUCEI 80S RIBOSOME / タイプ: RIBOSOME / 詳細: MICROGRAPHS SELECTED VISUALLY AFTER SCANNING |
|---|---|
| 緩衝液 | 名称: 20 MM HEPES, 10MM MGCL2, 500 MM KCL AND 5 MM BETA-ME pH: 7.2 詳細: 20 MM HEPES, 10MM MGCL2, 500 MM KCL AND 5 MM BETA-ME |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | 詳細: CARBON |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 詳細: LIQUID ETHAN |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai F30 / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TECNAI F30 / 日付: 2011年1月1日 詳細: MICROGRAPHS SCANNED WITH NIKON SUPER COOLSCAN 9000ED |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 59000 X / 最大 デフォーカス(公称値): 4000 nm / 最小 デフォーカス(公称値): 1500 nm / Cs: 2 mm |
| 試料ホルダ | 温度: 93.15 K |
| 撮影 | 電子線照射量: 25 e/Å2 / フィルム・検出器のモデル: KODAK SO-163 FILM |
| 画像スキャン | デジタル画像の数: 1102 |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: PHASE-FLIPPING | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 手法: REFERENCE BASED RECONSTRUCTION METHOD / 解像度: 5.57 Å / 粒子像の数: 164000 / ピクセルサイズ(公称値): 1.09 Å 詳細: SUBMISSION BASED ON EXPERIMENTAL DATA FROM EMDB EMD-2239 (DEPOSITION ID: 11259). THE STRUCTURE OF THE 80S FROM YEAST (3U5B AND OTHERS) AS WELL AS THE STRUCTURE OF THE 60S FROM TETRAHYMENA ...詳細: SUBMISSION BASED ON EXPERIMENTAL DATA FROM EMDB EMD-2239 (DEPOSITION ID: 11259). THE STRUCTURE OF THE 80S FROM YEAST (3U5B AND OTHERS) AS WELL AS THE STRUCTURE OF THE 60S FROM TETRAHYMENA THERMOPHILA (4A17 AND OTHERS) WERE USED AS STARTING MODEL FOR THE 60S SUBUNIT MODEL. THE 40S FROM TETRAHYMENA THERMOPHILA (2XZM AND 2XZN) AS WELL AS THE 80S FROM YEAST WERE USED AS STARTING MODEL FOR THE 40S SUBUNIT MODEL. THE 80S MODEL OF TRITICUM AESTIVUM (3IZR AND OTHERS) WAS USED TO FIT MISSING PROTEINS FORM THE TWO X-RAY STRUCTURES. 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: REAL / 詳細: REFINEMENT PROTOCOL--RIGID BODY | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 |
|
