 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 4v6m | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Structure of the ribosome-SecYE complex in the membrane environment | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | RIBOSOME/RIBOSOMAL PROTEIN / RIBOSOMAL PROTEIN / RIBONUCLEOPROTEIN / NUCLEOTIDE-BINDING / PROTEIN BIOSYNTHESIS / TRANSLATION / ZINC-FINGER / 70S RIBOSOME / RIBOSOME / TRANSLOCON / SECYEG / NANODISC / RIBOSOME-RIBOSOMAL PROTEIN complex | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報cell septum assembly / Defective ABCA1 causes TGD / high-density lipoprotein particle receptor binding / peptidyl-methionine modification / HDL clearance / spherical high-density lipoprotein particle / Scavenging by Class B Receptors / negative regulation of response to cytokine stimulus / protein oxidation / regulation of intestinal cholesterol absorption ...cell septum assembly / Defective ABCA1 causes TGD / high-density lipoprotein particle receptor binding / peptidyl-methionine modification / HDL clearance / spherical high-density lipoprotein particle / Scavenging by Class B Receptors / negative regulation of response to cytokine stimulus / protein oxidation / regulation of intestinal cholesterol absorption / vitamin transport / blood vessel endothelial cell migration / cholesterol import / high-density lipoprotein particle binding / negative regulation of heterotypic cell-cell adhesion / ABC transporters in lipid homeostasis / apolipoprotein A-I receptor binding / intracellular protein transmembrane transport / apolipoprotein receptor binding / negative regulation of cell adhesion molecule production / negative regulation of cytokine production involved in immune response / protein transport by the Sec complex / HDL assembly / negative regulation of very-low-density lipoprotein particle remodeling / phosphatidylcholine biosynthetic process / glucocorticoid metabolic process / protein-transporting ATPase activity / acylglycerol homeostasis / phosphatidylcholine-sterol O-acyltransferase activator activity / positive regulation of phospholipid efflux / Chylomicron remodeling / cellular response to lipoprotein particle stimulus / Chylomicron assembly / high-density lipoprotein particle clearance / phospholipid efflux / chylomicron / high-density lipoprotein particle remodeling / positive regulation of cholesterol metabolic process / lipid storage / reverse cholesterol transport / phospholipid homeostasis / high-density lipoprotein particle assembly / chemorepellent activity / low-density lipoprotein particle / lipoprotein biosynthetic process / cholesterol transfer activity / cholesterol transport / high-density lipoprotein particle / very-low-density lipoprotein particle / FtsZ-dependent cytokinesis / endothelial cell proliferation / regulation of Cdc42 protein signal transduction / HDL remodeling / cholesterol efflux / triglyceride homeostasis / Scavenging by Class A Receptors / negative regulation of interleukin-1 beta production / adrenal gland development / negative chemotaxis / cholesterol binding / cell division site / cholesterol biosynthetic process / positive regulation of Rho protein signal transduction / amyloid-beta formation / stringent response / ornithine decarboxylase inhibitor activity / transcription antitermination factor activity, RNA binding / protein secretion / misfolded RNA binding / protein transmembrane transporter activity / Group I intron splicing / positive regulation of cholesterol efflux / endocytic vesicle / RNA folding / negative regulation of tumor necrosis factor-mediated signaling pathway / membrane => GO:0016020 / transcriptional attenuation / protein targeting / endoribonuclease inhibitor activity / positive regulation of ribosome biogenesis / Scavenging of heme from plasma / RNA-binding transcription regulator activity / Retinoid metabolism and transport / cholesterol metabolic process / translational termination / negative regulation of cytoplasmic translation / four-way junction DNA binding / positive regulation of stress fiber assembly / DnaA-L2 complex / translation repressor activity / heat shock protein binding / negative regulation of translational initiation / regulation of mRNA stability / endocytic vesicle lumen / negative regulation of DNA-templated DNA replication initiation / positive regulation of substrate adhesion-dependent cell spreading / positive regulation of phagocytosis / mRNA regulatory element binding translation repressor activity / assembly of large subunit precursor of preribosome / cholesterol homeostasis 類似検索 - 分子機能 | |||||||||
| 生物種 |   Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 7.1 Å | |||||||||
 データ登録者 データ登録者 | Frauenfeld, J. / Gumbart, J. / van der Sluis, E.O. / Funes, S. / Gartmann, M. / Beatrix, B. / Mielke, T. / Berninghausen, O. / Becker, T. / Schulten, K. / Beckmann, R. | |||||||||
 引用 引用 | ジャーナル: Nat Struct Mol Biol / 年: 2011 タイトル: Cryo-EM structure of the ribosome-SecYE complex in the membrane environment. 著者: Jens Frauenfeld / James Gumbart / Eli O van der Sluis / Soledad Funes / Marco Gartmann / Birgitta Beatrix / Thorsten Mielke / Otto Berninghausen / Thomas Becker / Klaus Schulten / Roland Beckmann /  要旨: The ubiquitous SecY-Sec61 complex translocates nascent secretory proteins across cellular membranes and integrates membrane proteins into lipid bilayers. Several structures of mostly detergent- ...The ubiquitous SecY-Sec61 complex translocates nascent secretory proteins across cellular membranes and integrates membrane proteins into lipid bilayers. Several structures of mostly detergent-solubilized Sec complexes have been reported. Here we present a single-particle cryo-EM structure of the SecYEG complex in a membrane environment, bound to a translating ribosome, at subnanometer resolution. Using the SecYEG complex reconstituted in a so-called Nanodisc, we could trace the nascent polypeptide chain from the peptidyltransferase center into the membrane. The reconstruction allowed for the identification of ribosome-lipid interactions. The rRNA helix 59 (H59) directly contacts the lipid surface and appears to modulate the membrane in immediate vicinity to the proposed lateral gate of the protein-conducting channel (PCC). On the basis of our map and molecular dynamics simulations, we present a model of a signal anchor-gated PCC in the membrane. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 4v6m.cif.gz | 3.6 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb4v6m.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 4v6m.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 4v6m_validation.pdf.gz | 3.8 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 4v6m_full_validation.pdf.gz | 5.2 MB | 表示 | |
| XML形式データ | 4v6m_validation.xml.gz | 356.8 KB | 表示 | |
| CIF形式データ | 4v6m_validation.cif.gz | 580.7 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/v6/4v6mftp://data.pdbj.org/pub/pdb/validation_reports/v6/4v6m | HTTPS FTP |
-関連構造データ


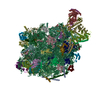


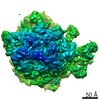
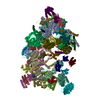
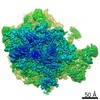
-リンク
 PDBj
PDBj




































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 5種, 5分子 AAAXAVB7B8
| #1: RNA鎖 | 分子量: 499690.031 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: RNA鎖 | 分子量: 3442.106 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: RNA鎖 | 分子量: 24876.777 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #26: RNA鎖 | 分子量: 38790.090 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #27: RNA鎖 | 分子量: 941612.375 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 2種, 3分子 AZA0A1
| #4: タンパク質 | 分子量: 11085.822 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #5: タンパク質 | 分子量: 23309.361 Da / 分子数: 2 / 由来タイプ: 合成 / 詳細: nanodiscs derived from human Apo-A1 / 由来: (合成) Homo sapiens (ヒト) / 参照: UniProt: P02647 |
-30S ribosomal protein ... , 20種, 20分子 ABACADAEAFAGAHAIAJAKALAMANAOAPAQARASATAU
| #6: タンパク質 | 分子量: 26650.475 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #7: タンパク質 | 分子量: 25900.117 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #8: タンパク質 | 分子量: 23383.002 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #9: タンパク質 | 分子量: 17498.203 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #10: タンパク質 | 分子量: 15727.512 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #11: タンパク質 | 分子量: 19923.959 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #12: タンパク質 | 分子量: 14015.361 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #13: タンパク質 | 分子量: 14755.074 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #14: タンパク質 | 分子量: 11755.597 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #15: タンパク質 | 分子量: 13739.778 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #16: タンパク質 | 分子量: 13636.961 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #17: タンパク質 | 分子量: 12997.271 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #18: タンパク質 | 分子量: 11475.364 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #19: タンパク質 | 分子量: 10188.687 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #20: タンパク質 | 分子量: 9207.572 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #21: タンパク質 | 分子量: 9593.296 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #22: タンパク質 | 分子量: 8874.276 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #23: タンパク質 | 分子量: 10324.160 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #24: タンパク質 | 分子量: 9577.268 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #25: タンパク質 | 分子量: 8392.844 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-Preprotein translocase ... , 2種, 2分子 BABB
| #28: タンパク質 | 分子量: 47669.277 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #29: タンパク質 | 分子量: 12623.296 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+50S ribosomal protein ... , 30種, 30分子 B5B6BDBEBFBGBHBIBJBKBLBMBNBOBPBQBRBSBTBUBVBWBXBYBZB0B1B2B3B4
-非ポリマー , 2種, 133分子 

| #60: 化合物 | ChemComp-PEV / (  分子量: 720.012 Da / 分子数: 101 / 由来タイプ: 合成 / 式: C39H78NO8P / コメント: POPE, リン脂質*YM 分子量: 720.012 Da / 分子数: 101 / 由来タイプ: 合成 / 式: C39H78NO8P / コメント: POPE, リン脂質*YM#61: 化合物 | ChemComp-PGV / (  分子量: 749.007 Da / 分子数: 32 / 由来タイプ: 合成 / 式: C40H77O10P / コメント: リン脂質*YM 分子量: 749.007 Da / 分子数: 32 / 由来タイプ: 合成 / 式: C40H77O10P / コメント: リン脂質*YM |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: An active E. coli SecYEG complex embedded in a lipid bilayer (Nanodisc), bound to a translating E. coli ribosome タイプ: RIBOSOME 詳細: The heterotrimeric SecYEG complex was embedded in a lipid bilayer (nascent HDL, Nanodisc) |
|---|---|
| 緩衝液 | 名称: 20 mM Hepes (pH 7.2), 100 mM KOAc, 10 mM Mg(OAc)2, 1 mM DTT, 250 microg/ml chloramphenicol pH: 7.2 詳細: 20 mM Hepes (pH 7.2), 100 mM KOAc, 10 mM Mg(OAc)2, 1 mM DTT, 250 microg/ml chloramphenicol |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE / 詳細: liquid ethane was used as a cryogen |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI POLARA 300 |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 39000 X / 倍率(補正後): 38000 X / 最大 デフォーカス(公称値): 4500 nm / 最小 デフォーカス(公称値): 1000 nm / Cs: 2.26 mm |
| 撮影 | 電子線照射量: 22 e/Å2 / フィルム・検出器のモデル: KODAK SO-163 FILM |
| 放射 | プロトコル: SINGLE WAVELENGTH / 単色(M)・ラウエ(L): M / 散乱光タイプ: x-ray |
| 放射波長 | 相対比: 1 |
- 解析
解析
| EMソフトウェア |
| ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: DEFOCUS GROUP VOLUMES | ||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||
| 3次元再構成 | 解像度: 7.1 Å / 粒子像の数: 85664 / 対称性のタイプ: POINT | ||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: REAL / 詳細: REFINEMENT PROTOCOL--flexible fitting | ||||||||||||
| 精密化ステップ | サイクル: LAST
|
