Mitotic Telophase/Cytokinesis / regulation of protein localization to cell cortex / Mitotic Metaphase/Anaphase Transition / synaptonemal complex disassembly / Activation of NIMA Kinases NEK9, NEK6, NEK7 / homologous chromosome segregation / polo kinase / mitotic nuclear membrane disassembly / protein localization to nuclear envelope / Phosphorylation of Emi1 ...Mitotic Telophase/Cytokinesis / regulation of protein localization to cell cortex / Mitotic Metaphase/Anaphase Transition / synaptonemal complex disassembly / Activation of NIMA Kinases NEK9, NEK6, NEK7 / homologous chromosome segregation / polo kinase / mitotic nuclear membrane disassembly / protein localization to nuclear envelope / Phosphorylation of Emi1 / metaphase/anaphase transition of mitotic cell cycle / nuclear membrane disassembly / synaptonemal complex / female meiosis chromosome segregation / Golgi inheritance / Phosphorylation of the APC/C / anaphase-promoting complex binding / outer kinetochore / positive regulation of ubiquitin protein ligase activity / microtubule bundle formation / double-strand break repair via alternative nonhomologous end joining / mitotic chromosome condensation / Polo-like kinase mediated events / regulation of mitotic spindle assembly / Golgi Cisternae Pericentriolar Stack Reorganization / centrosome cycle / regulation of mitotic metaphase/anaphase transition / positive regulation of ubiquitin-protein transferase activity / sister chromatid cohesion / regulation of mitotic cell cycle phase transition / mitotic spindle assembly checkpoint signaling / mitotic spindle pole / spindle midzone / mitotic G2 DNA damage checkpoint signaling / regulation of anaphase-promoting complex-dependent catabolic process / mitotic cytokinesis / establishment of mitotic spindle orientation / mitotic sister chromatid segregation / positive regulation of proteolysis / negative regulation of double-strand break repair via homologous recombination / Regulation of MITF-M-dependent genes involved in cell cycle and proliferation / Cyclin A/B1/B2 associated events during G2/M transition / protein localization to chromatin / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / centriole / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / Loss of Nlp from mitotic centrosomes / Loss of proteins required for interphase microtubule organization from the centrosome / Recruitment of mitotic centrosome proteins and complexes / Recruitment of NuMA to mitotic centrosomes / Anchoring of the basal body to the plasma membrane / regulation of mitotic cell cycle / Resolution of Sister Chromatid Cohesion / Condensation of Prophase Chromosomes / regulation of cytokinesis / AURKA Activation by TPX2 / mitotic spindle organization / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / establishment of protein localization / RHO GTPases Activate Formins / protein destabilization / peptidyl-serine phosphorylation / kinetochore / centriolar satellite / positive regulation of protein localization to nucleus / spindle / G2/M transition of mitotic cell cycle / The role of GTSE1 in G2/M progression after G2 checkpoint / Separation of Sister Chromatids / spindle pole / Regulation of PLK1 Activity at G2/M Transition / positive regulation of proteasomal ubiquitin-dependent protein catabolic process / double-strand break repair / mitotic cell cycle / microtubule cytoskeleton / midbody / microtubule binding / protein phosphorylation / protein ubiquitination / protein kinase activity / regulation of cell cycle / protein serine kinase activity / protein serine/threonine kinase activity / centrosome / protein kinase binding / negative regulation of apoptotic process / chromatin / magnesium ion binding / negative regulation of transcription by RNA polymerase II / nucleoplasm / ATP binding / identical protein binding / nucleus / cytosol / cytoplasm 類似検索 - 分子機能
POLO box domain / Polo-like kinase 1, catalytic domain / Second polo-box domain / First polo-box domain / POLO box domain superfamily / POLO box duplicated region / POLO box domain / POLO box domain profile. / Arylsulfatase, C-terminal domain / Serine/threonine-protein kinase, active site ...POLO box domain / Polo-like kinase 1, catalytic domain / Second polo-box domain / First polo-box domain / POLO box domain superfamily / POLO box duplicated region / POLO box domain / POLO box domain profile. / Arylsulfatase, C-terminal domain / Serine/threonine-protein kinase, active site / Serine/Threonine protein kinases active-site signature. / Protein kinase domain / Serine/Threonine protein kinases, catalytic domain / Protein kinase, ATP binding site / Protein kinases ATP-binding region signature. / Protein kinase domain profile. / Protein kinase domain / Protein kinase-like domain superfamily / 2-Layer Sandwich / Alpha Beta 類似検索 - ドメイン・相同性
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 要素
要素 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) X線回折 /
X線回折 /  データ登録者
データ登録者 引用
引用 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード


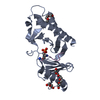



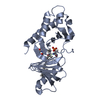


 PDBj
PDBj







 集合体
集合体


 タイプ: Polypeptide / クラス: 酵素阻害剤 / 分子量: 935.056 Da / 分子数: 1 / 由来タイプ: 合成 / 参照: Phosphothreonine ester-containing peptide
タイプ: Polypeptide / クラス: 酵素阻害剤 / 分子量: 935.056 Da / 分子数: 1 / 由来タイプ: 合成 / 参照: Phosphothreonine ester-containing peptide 分子量: 18.015 Da / 分子数: 218 / 由来タイプ: 天然 / 式: H2O
分子量: 18.015 Da / 分子数: 218 / 由来タイプ: 天然 / 式: H2O 試料調製
試料調製 解析
解析