Journal: Structure / Year: 2013 Title: Structural insights into the intrinsic self-assembly of Par-3 N-terminal domain. Authors: Yan Zhang / Wenjuan Wang / Jia Chen / Kai Zhang / Feng Gao / Bingquan Gao / Shuai Zhang / Mingdong Dong / Flemming Besenbacher / Weimin Gong / Mingjie Zhang / Fei Sun / Wei Feng / Abstract: Par-3, the central organizer of the Par-3/Par-6/atypical protein kinase C complex, is a multimodular scaffold protein that is essential for cell polarity establishment and maintenance. The N-terminal ...Par-3, the central organizer of the Par-3/Par-6/atypical protein kinase C complex, is a multimodular scaffold protein that is essential for cell polarity establishment and maintenance. The N-terminal domain (NTD) of Par-3 is capable of self-association to form filament-like structures, although the underlying mechanism is poorly understood. Here, we determined the crystal structure of Par-3 NTD and solved the filament structure by cryoelectron microscopy. We found that an intrinsic "front-to-back" interaction mode is important for Par-3 NTD self-association and that both the lateral and longitudinal packing within the filament are mediated by electrostatic interactions. Disruptions of the lateral or longitudinal packing significantly impaired Par-3 NTD self-association and thereby impacted the Par-3-mediated epithelial polarization. We finally demonstrated that a Par-3 NTD-like domain from histidine ammonia-lyase also harbors a similar self-association capacity. This work unequivocally provides the structural basis for Par-3 NTD self-association and characterizes one type of protein domain that can self-assemble via electrostatic interactions.
This protein forms helical filament in solution. Cryo-electron microscopic reconstruction revealed that the dimer in the crystallographic asymmetric unit is the same as the building block of the helical filament.
Resolution: 2.9→39.94 Å / Cor.coef. Fo:Fc: 0.918 / Cor.coef. Fo:Fc free: 0.86 / SU B: 19.255 / SU ML: 0.35 / Cross valid method: THROUGHOUT / σ(F): 0 / ESU R: 1.008 / ESU R Free: 0.415 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN USED IF PRESENT IN THE INPUT
Rfactor
Num. reflection
% reflection
Selection details
Rfree
0.29361
301
4.7 %
RANDOM
Rwork
0.23005
-
-
-
obs
0.23296
6058
96.33 %
-
all
-
6289
-
-
Solvent computation
Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å / Solvent model: MASK
Displacement parameters
Biso mean: 55.809 Å2
Baniso -1
Baniso -2
Baniso -3
1-
-0.43 Å2
0 Å2
0 Å2
2-
-
-0.43 Å2
0 Å2
3-
-
-
0.85 Å2
Refinement step
Cycle: LAST / Resolution: 2.9→39.94 Å
Protein
Nucleic acid
Ligand
Solvent
Total
Num. atoms
1323
0
0
34
1357
Refine LS restraints
Refine-ID
Type
Dev ideal
Dev ideal target
Number
X-RAY DIFFRACTION
r_bond_refined_d
0.012
0.019
1345
X-RAY DIFFRACTION
r_angle_refined_deg
1.791
1.948
1816
X-RAY DIFFRACTION
r_dihedral_angle_1_deg
7.769
5
164
X-RAY DIFFRACTION
r_dihedral_angle_2_deg
34.93
23.333
69
X-RAY DIFFRACTION
r_dihedral_angle_3_deg
19.812
15
235
X-RAY DIFFRACTION
r_dihedral_angle_4_deg
17.571
15
14
X-RAY DIFFRACTION
r_chiral_restr
0.12
0.2
204
X-RAY DIFFRACTION
r_gen_planes_refined
0.006
0.02
1022
LS refinement shell
Resolution: 2.9→2.975 Å / Total num. of bins used: 20
Rfactor
Num. reflection
% reflection
Rfree
0.325
21
-
Rwork
0.307
343
-
obs
-
343
89.88 %
+
About Yorodumi
-
News
-
Feb 9, 2022. New format data for meta-information of EMDB entries
New format data for meta-information of EMDB entries
Version 3 of the EMDB header file is now the official format.
The previous official version 1.9 will be removed from the archive.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information
 X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation
 Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads



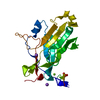
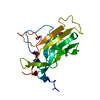




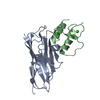


 PDBj
PDBj Assembly
Assembly



 Mass: 18.015 Da / Num. of mol.: 34 / Source method: isolated from a natural source / Formula: H2O
Mass: 18.015 Da / Num. of mol.: 34 / Source method: isolated from a natural source / Formula: H2O Sample preparation
Sample preparation Processing
Processing