 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-3qhe: Crystal structure of the complex between the armadillo repeat dom... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 3qhe | ||||||
|---|---|---|---|---|---|---|---|
| Title | Crystal structure of the complex between the armadillo repeat domain of adenomatous polyposis coli and the tyrosine-rich domain of Sam68 | ||||||
 Components Components |
| ||||||
 Keywords Keywords | signaling protein/splicing / Armadillo repeat superhelix / Regulation of Wnt signaling / tumor suppressor protein / adaptor protein / RNA-binding protein / signaling protein-splicing protein complex / Structural Genomics / NPPSFA / National Project on Protein Structural and Functional Analyses / RIKEN Structural Genomics/Proteomics Initiative / RSGI / signaling protein-splicing complex | ||||||
| Function / homology |  Function and homology information Function and homology informationregulation of RNA export from nucleus / Grb2-Sos complex / APC truncation mutants are not K63 polyubiquitinated / PTK6 Regulates Proteins Involved in RNA Processing / positive regulation of RNA export from nucleus / negative regulation of cell cycle G1/S phase transition / gamma-catenin binding / negative regulation of cyclin-dependent protein serine/threonine kinase activity / regulation of microtubule-based movement / regulation of attachment of spindle microtubules to kinetochore ...regulation of RNA export from nucleus / Grb2-Sos complex / APC truncation mutants are not K63 polyubiquitinated / PTK6 Regulates Proteins Involved in RNA Processing / positive regulation of RNA export from nucleus / negative regulation of cell cycle G1/S phase transition / gamma-catenin binding / negative regulation of cyclin-dependent protein serine/threonine kinase activity / regulation of microtubule-based movement / regulation of attachment of spindle microtubules to kinetochore / positive regulation of pseudopodium assembly / positive regulation of protein localization to centrosome / bicellular tight junction assembly / pattern specification process / negative regulation of microtubule depolymerization / catenin complex / beta-catenin destruction complex / heart valve development / APC truncation mutants have impaired AXIN binding / AXIN missense mutants destabilize the destruction complex / Truncations of AMER1 destabilize the destruction complex / poly(A) binding / regulation of microtubule-based process / microtubule plus-end binding / Beta-catenin phosphorylation cascade / Signaling by GSK3beta mutants / CTNNB1 S33 mutants aren't phosphorylated / CTNNB1 S37 mutants aren't phosphorylated / CTNNB1 S45 mutants aren't phosphorylated / CTNNB1 T41 mutants aren't phosphorylated / protein kinase regulator activity / Wnt signalosome / cell fate specification / Disassembly of the destruction complex and recruitment of AXIN to the membrane / poly(U) RNA binding / endocardial cushion morphogenesis / molecular function inhibitor activity / mitotic spindle assembly checkpoint signaling / negative regulation of G1/S transition of mitotic cell cycle / Apoptotic cleavage of cellular proteins / regulation of alternative mRNA splicing, via spliceosome / dynein complex binding / regulation of RNA splicing / mitotic cytokinesis / positive regulation of translational initiation / lateral plasma membrane / bicellular tight junction / signaling adaptor activity / SH2 domain binding / Deactivation of the beta-catenin transactivating complex / protein tyrosine kinase binding / adherens junction / Degradation of beta-catenin by the destruction complex / G1/S transition of mitotic cell cycle / negative regulation of canonical Wnt signaling pathway / beta-catenin binding / kinetochore / SH3 domain binding / G2/M transition of mitotic cell cycle / ruffle membrane / Wnt signaling pathway / Ovarian tumor domain proteases / mRNA processing / positive regulation of protein catabolic process / insulin receptor signaling pathway / cell migration / nervous system development / lamellipodium / T cell receptor signaling pathway / positive regulation of cold-induced thermogenesis / protein-containing complex assembly / spermatogenesis / regulation of apoptotic process / microtubule binding / proteasome-mediated ubiquitin-dependent protein catabolic process / microtubule / regulation of cell cycle / cell adhesion / positive regulation of cell migration / positive regulation of apoptotic process / protein domain specific binding / negative regulation of cell population proliferation / negative regulation of DNA-templated transcription / mRNA binding / ubiquitin protein ligase binding / centrosome / DNA damage response / protein kinase binding / protein-containing complex binding / perinuclear region of cytoplasm / negative regulation of transcription by RNA polymerase II / Golgi apparatus / protein-containing complex / DNA binding / RNA binding / nucleoplasm / identical protein binding / nucleus / membrane / plasma membrane Similarity search - Function | ||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.4 Å X-RAY DIFFRACTION / SYNCHROTRON / MOLECULAR REPLACEMENT / Resolution: 2.4 Å | ||||||
 Authors Authors | Morishita, E.C.J. / Murayama, K. / Kato-Murayama, M. / Ishizuku-Katsura, Y. / Tomabechi, Y. / Terada, T. / Handa, N. / Shirouzu, M. / Akiyama, T. / Yokoyama, S. / RIKEN Structural Genomics/Proteomics Initiative (RSGI) | ||||||
 Citation Citation | Journal: Structure / Year: 2011 Title: Crystal structures of the armadillo repeat domain of adenomatous polyposis coli and its complex with the tyrosine-rich domain of sam68 Authors: Morishita, E.C. / Murayama, K. / Kato-Murayama, M. / Ishizuka-Katsura, Y. / Tomabechi, Y. / Hayashi, T. / Terada, T. / Handa, N. / Shirouzu, M. / Akiyama, T. / Yokoyama, S. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 3qhe.cif.gz | 270.5 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb3qhe.ent.gz | 222.2 KB | Display | PDB format |
| PDBx/mmJSON format | 3qhe.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 3qhe_validation.pdf.gz | 449.6 KB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 3qhe_full_validation.pdf.gz | 455.2 KB | Display | |
| Data in XML | 3qhe_validation.xml.gz | 26.8 KB | Display | |
| Data in CIF | 3qhe_validation.cif.gz | 38.6 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/qh/3qheftp://data.pdbj.org/pub/pdb/validation_reports/qh/3qhe | HTTPS FTP |
-Related structure data
| Related structure data | 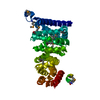 3au3SC S: Starting model for refinement C: citing same article ( |
|---|---|
| Similar structure data |






-Links
 PDBj
PDBj




- Assembly
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 37502.219 Da / Num. of mol.: 2 / Fragment: Armadillo repeat domain, residues 396-732 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Description: E. coli cell-free system / Plasmid: pCR2.1 / Production host: cell free synthsis (unknown) / References: UniProt: P25054#2: Protein | Mass: 6493.303 Da / Num. of mol.: 2 / Fragment: Tyrosine-rich domain, residues 365-419 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Description: E. coli cell-free system / Plasmid: pCR2.1 / Production host: cell free synthsis (unknown) / References: UniProt: Q07666#3: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 262 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 262 / Source method: isolated from a natural source / Formula: H2O |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.28 Å3/Da / Density % sol: 46.15 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, sitting drop / pH: 5.5 Details: 100mM BIS-TRIS (pH 5.5), 150mM NaCl, 20% PEG 3350, VAPOR DIFFUSION, SITTING DROP, temperature 293K |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: SLS  / Beamline: X06DA / Wavelength: 1 Å / Beamline: X06DA / Wavelength: 1 Å |
| Detector | Type: PSI PILATUS 6M / Detector: PIXEL / Date: Mar 30, 2010 |
| Radiation | Monochromator: GRAPHITE / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 1 Å / Relative weight: 1 |
| Reflection | Resolution: 2.4→64.18 Å / Num. all: 31344 / Num. obs: 31344 / % possible obs: 100 % / Observed criterion σ(F): 0 / Observed criterion σ(I): -3 / Redundancy: 11.2 % / Rmerge(I) obs: 0.095 / Rsym value: 0.091 / Net I/σ(I): 7.1 |
| Reflection shell | Resolution: 2.4→2.53 Å / Redundancy: 10.8 % / Rmerge(I) obs: 0.388 / Mean I/σ(I) obs: 2 / Num. unique all: 4539 / Rsym value: 0.37 / % possible all: 100 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MOLECULAR REPLACEMENT Starting model: PDB ENTRY 3AU3 Resolution: 2.4→64.18 Å / Cor.coef. Fo:Fc: 0.938 / Cor.coef. Fo:Fc free: 0.908 / SU B: 14.657 / SU ML: 0.157 / Cross valid method: THROUGHOUT / ESU R Free: 0.239 / Stereochemistry target values: MAXIMUM LIKELIHOOD Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS; ASN709-LEU710 and LEU710-ILE711 C-N bonds in chain C, 0.91A and 1.05A, respectively, lie outside of the accepted range for the peptide bond ...Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS; ASN709-LEU710 and LEU710-ILE711 C-N bonds in chain C, 0.91A and 1.05A, respectively, lie outside of the accepted range for the peptide bond (1.30-1.45). depositors said that it was hard to build them suitably because these residues are in a region of poor density.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Solvent computation | Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.4 Å / Solvent model: MASK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 32.716 Å2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 2.4→64.18 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Resolution: 2.4→2.462 Å / Total num. of bins used: 20
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|
