 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 3j92 | ||||||
|---|---|---|---|---|---|---|---|
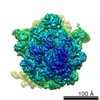
| タイトル | Structure and assembly pathway of the ribosome quality control complex | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | RIBOSOME/LIGASE / RWD / RING / quality control / RIBOSOME-LIGASE complex | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報alpha-aminoacyl-tRNA binding / CAT tailing / RQC complex / nuclear export / ribosome-associated ubiquitin-dependent protein catabolic process / ribosomal large subunit binding / protein autoubiquitination / cytosolic ribosome / rescue of stalled ribosome / RING-type E3 ubiquitin transferase ...alpha-aminoacyl-tRNA binding / CAT tailing / RQC complex / nuclear export / ribosome-associated ubiquitin-dependent protein catabolic process / ribosomal large subunit binding / protein autoubiquitination / cytosolic ribosome / rescue of stalled ribosome / RING-type E3 ubiquitin transferase / Antigen processing: Ubiquitination & Proteasome degradation / ubiquitin protein ligase activity / protein-containing complex assembly / tRNA binding / zinc ion binding / nucleus / cytosol 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.6 Å | ||||||
 データ登録者 データ登録者 | Shao, S. / Brown, A. / Santhanam, B. / Hegde, R.S. | ||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2015 タイトル: Structure and assembly pathway of the ribosome quality control complex. 著者: Sichen Shao / Alan Brown / Balaji Santhanam / Ramanujan S Hegde /  要旨: During ribosome-associated quality control, stalled ribosomes are split into subunits and the 60S-housed nascent polypeptides are poly-ubiquitinated by Listerin. How this low-abundance ubiquitin ...During ribosome-associated quality control, stalled ribosomes are split into subunits and the 60S-housed nascent polypeptides are poly-ubiquitinated by Listerin. How this low-abundance ubiquitin ligase targets rare stall-generated 60S among numerous empty 60S is unknown. Here, we show that Listerin specificity for nascent chain-60S complexes depends on nuclear export mediator factor (NEMF). The 3.6 Å cryo-EM structure of a nascent chain-containing 60S-Listerin-NEMF complex revealed that NEMF makes multiple simultaneous contacts with 60S and peptidyl-tRNA to sense nascent chain occupancy. Structural and mutational analyses showed that ribosome-bound NEMF recruits and stabilizes Listerin's N-terminal domain, while Listerin's C-terminal RWD domain directly contacts the ribosome to position the adjacent ligase domain near the nascent polypeptide exit tunnel. Thus, highly specific nascent chain targeting by Listerin is imparted by the avidity gained from a multivalent network of context-specific individually weak interactions, highlighting a new principle of client recognition during protein quality control. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
 UCSF Chimera
UCSF Chimera- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3j92.cif.gz | 3.6 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3j92.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 3j92.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 3j92_validation.pdf.gz | 1.3 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 3j92_full_validation.pdf.gz | 1.5 MB | 表示 | |
| XML形式データ | 3j92_validation.xml.gz | 228.7 KB | 表示 | |
| CIF形式データ | 3j92_validation.cif.gz | 402.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/j9/3j92ftp://data.pdbj.org/pub/pdb/validation_reports/j9/3j92 | HTTPS FTP |
-関連構造データ



-リンク
 PDBj
PDBj





























- 集合体
集合体
| 登録構造単位 | 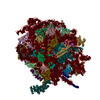
|
|---|---|
| 1 |
|
-要素
+タンパク質 , 47種, 51分子 ABCDEFGHIJLMNOPQRSTUVWXYZabcde...
-RNA鎖 , 4種, 4分子 2578
| #48: RNA鎖 | 分子量: 24816.811 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #49: RNA鎖 | 分子量: 1187230.000 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #50: RNA鎖 | 分子量: 38691.914 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #51: RNA鎖 | 分子量: 50143.648 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-非ポリマー , 2種, 164分子 

| #52: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 159 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 159 / 由来タイプ: 合成 / 式: Mg#53: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 5 / 由来タイプ: 合成 / 式: Zn |
|---|
-詳細
| Has protein modification | Y |
|---|---|
| 配列の詳細 | ALTHOUGH ONLY ONE INSTANCE EACH OF PROTEINS NEMF AND LISTERIN IS PRESENT WITHIN THE COMPLEX, EACH ...ALTHOUGH ONLY ONE INSTANCE EACH OF PROTEINS NEMF AND LISTERIN IS PRESENT WITHIN THE COMPLEX, EACH OF THESE PROTEINS IS REPRESENTE |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 緩衝液 | 名称: 50 mM HEPES, 100 mM potassium acetate, 5 mM magnesium acetate, 1 mM DTT pH: 7.4 詳細: 50 mM HEPES, 100 mM potassium acetate, 5 mM magnesium acetate, 1 mM DTT | ||||||||||||||||||||||||||||
| 試料 | 濃度: 0.3 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||
| 試料支持 | 詳細: Quantifoil R2/2 on 400 mesh Cu grid with thin carbon support | ||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK I / 凍結剤: ETHANE / 湿度: 100 % 詳細: Blot for 3 seconds before plunging into liquid ethane (FEI VITROBOT MARK I). 手法: Blot for 3 seconds before plunging |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company | ||||||
|---|---|---|---|---|---|---|---|
| EM imaging | 加速電圧: 300 kV / 倍率(補正後): 104478 X / 詳細: Objective lens astigmatism was corrected at 59,000 times magnification. / 電子線源:
| ||||||
| 撮影 | 電子線照射量: 30 e/Å2 フィルム・検出器のモデル: FEI FALCON II (4k x 4k) |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0091 2014/10/05 / 分類: 精密化 / Contact author: Garib N. Murshudov / Contact author email: garib[at]mrc-lmb.cam.ac.uk 解説: (un)restrained refinement or idealisation of macromolecular structures | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| |||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | |||||||||||||||||||||
| 3次元再構成 | 解像度: 3.6 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 63826 / ピクセルサイズ(公称値): 1.34 Å / ピクセルサイズ(実測値): 1.34 Å / 対称性のタイプ: POINT | |||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT / 空間: RECIPROCAL / 詳細: REFINEMENT PROTOCOL--rigid body | |||||||||||||||||||||
| 原子モデル構築 |
| |||||||||||||||||||||
| 精密化 | 詳細: Hydrogens have been added in their riding positions | |||||||||||||||||||||
| 精密化ステップ | サイクル: LAST
|

