 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
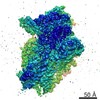
| 登録情報 | データベース: PDB / ID: 3j81 | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | CryoEM structure of a partial yeast 48S preinitiation complex | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | RIBOSOME / eukaryotic translation initiation / 48S / small ribosome subunit | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報formation of translation initiation ternary complex / Recycling of eIF2:GDP / Cellular response to mitochondrial stress / ABC-family proteins mediated transport / methionyl-initiator methionine tRNA binding / translation reinitiation / eukaryotic translation initiation factor 2 complex / formation of cytoplasmic translation initiation complex / multi-eIF complex / eukaryotic 43S preinitiation complex ...formation of translation initiation ternary complex / Recycling of eIF2:GDP / Cellular response to mitochondrial stress / ABC-family proteins mediated transport / methionyl-initiator methionine tRNA binding / translation reinitiation / eukaryotic translation initiation factor 2 complex / formation of cytoplasmic translation initiation complex / multi-eIF complex / eukaryotic 43S preinitiation complex / formation of translation preinitiation complex / eukaryotic 48S preinitiation complex / positive regulation of translational fidelity / protein-synthesizing GTPase / Formation of the ternary complex, and subsequently, the 43S complex / Translation initiation complex formation / Ribosomal scanning and start codon recognition / Formation of a pool of free 40S subunits / L13a-mediated translational silencing of Ceruloplasmin expression / ribosomal small subunit binding / 90S preribosome / regulation of translational fidelity / translation regulator activity / translation initiation factor binding / translation initiation factor activity / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / small-subunit processome / translational initiation / maintenance of translational fidelity / cytoplasmic stress granule / rRNA processing / double-stranded RNA binding / regulation of translation / ribosome binding / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / cytoplasmic translation / rRNA binding / structural constituent of ribosome / ribosome / translation / ribonucleoprotein complex / mRNA binding / GTPase activity / protein kinase binding / GTP binding / nucleolus / RNA binding / zinc ion binding / nucleus / cytosol / cytoplasm 類似検索 - 分子機能 | ||||||
| 生物種 |  Kluyveromyces lactis (酵母) Kluyveromyces lactis (酵母)synthetic construct (人工物) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4 Å | ||||||
 データ登録者 データ登録者 | Hussain, T. / Llacer, J.L. / Fernandez, I.S. / Savva, C.G. / Ramakrishnan, V. | ||||||
 引用 引用 | ジャーナル: Cell / 年: 2014 タイトル: Structural changes enable start codon recognition by the eukaryotic translation initiation complex. 著者: Tanweer Hussain / Jose L Llácer / Israel S Fernández / Antonio Munoz / Pilar Martin-Marcos / Christos G Savva / Jon R Lorsch / Alan G Hinnebusch / V Ramakrishnan /   要旨: During eukaryotic translation initiation, initiator tRNA does not insert fully into the P decoding site on the 40S ribosomal subunit. This conformation (POUT) is compatible with scanning mRNA for the ...During eukaryotic translation initiation, initiator tRNA does not insert fully into the P decoding site on the 40S ribosomal subunit. This conformation (POUT) is compatible with scanning mRNA for the AUG start codon. Base pairing with AUG is thought to promote isomerization to a more stable conformation (PIN) that arrests scanning and promotes dissociation of eIF1 from the 40S subunit. Here, we present a cryoEM reconstruction of a yeast preinitiation complex at 4.0 Å resolution with initiator tRNA in the PIN state, prior to eIF1 release. The structure reveals stabilization of the codon-anticodon duplex by the N-terminal tail of eIF1A, changes in the structure of eIF1 likely instrumental in its subsequent release, and changes in the conformation of eIF2. The mRNA traverses the entire mRNA cleft and makes connections to the regulatory domain of eIF2?, eIF1A, and ribosomal elements that allow recognition of context nucleotides surrounding the AUG codon. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 3j81.cif.gz | 2.1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb3j81.ent.gz | 1.6 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 3j81.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 3j81_validation.pdf.gz | 1.3 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 3j81_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 3j81_validation.xml.gz | 157.1 KB | 表示 | |
| CIF形式データ | 3j81_validation.cif.gz | 270.7 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/j8/3j81ftp://data.pdbj.org/pub/pdb/validation_reports/j8/3j81 | HTTPS FTP |
-関連構造データ

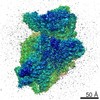


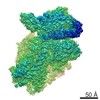
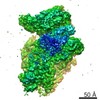
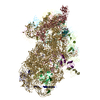



-リンク
 PDBj
PDBj































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-RNA鎖 , 3種, 3分子 213
| #1: RNA鎖 | 分子量: 579545.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Kluyveromyces lactis (酵母) |
|---|---|
| #36: RNA鎖 | 分子量: 24223.482 Da / 分子数: 1 / 由来タイプ: 合成 / 詳細: In vitro transcribed tRNA, aminoacylated |
| #37: RNA鎖 | 分子量: 7786.572 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
+タンパク質 , 38種, 38分子 ABCDEFGHIJKLMNOPQRSTUVWXYZabcf...
-タンパク質・ペプチド , 1種, 1分子 h
| #35: タンパク質・ペプチド | 分子量: 3354.243 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主:  |
|---|
-非ポリマー , 3種, 85分子 

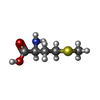
| #43: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 81 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 81 / 由来タイプ: 合成 / 式: Mg#44: 化合物 |  分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn#45: 化合物 | ChemComp-MET / |  タイプ: L-peptide linking / 分子量: 149.211 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C5H11NO2S タイプ: L-peptide linking / 分子量: 149.211 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C5H11NO2S |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 値: 1.2 MDa / 実験値: NO | ||||||||||||||||||||||||||||||||
| 緩衝液 | 名称: 20 mM MES-KOH, 40 mM potassium acetate, 10 mM ammonium acetate, 8 mM magnesium acetate, 2 mM DTT pH: 6.5 詳細: 20 mM MES-KOH, 40 mM potassium acetate, 10 mM ammonium acetate, 8 mM magnesium acetate, 2 mM DTT | ||||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.17 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||||
| 試料支持 | 詳細: Quantifoil R2/2 400 mesh copper grids with 4-5 nm thin carbon on top | ||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK I / 凍結剤: ETHANE / Temp: 120 K / 湿度: 100 % 詳細: Blot for 2.5 seconds before plunging into liquid ethane (FEI VITROBOT MARK I) 手法: Blot for 2.5 seconds before plunging |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI POLARA 300 / 日付: 2013年10月28日 詳細: Complete dataset was collected in 5 non-consecutive days |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 78000 X / 倍率(補正後): 104478 X / 最大 デフォーカス(公称値): 4000 nm / 最小 デフォーカス(公称値): 1600 nm / Cs: 2 mm |
| 試料ホルダ | 資料ホルダタイプ: GATAN LIQUID NITROGEN |
| 撮影 | 電子線照射量: 28 e/Å2 フィルム・検出器のモデル: FEI FALCON II (4k x 4k) |
| 画像スキャン | デジタル画像の数: 1791 |
| 放射波長 | 相対比: 1 |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0077 2014/05/16 / 分類: 精密化 / Contact author: Garib N. Murshudov / Contact author email: garib[at]mrc-lmb.cam.ac.uk 解説: (un)restrained refinement or idealisation of macromolecular structures | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||
| CTF補正 | 詳細: Each particle | ||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||
| 3次元再構成 | 解像度: 4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 29698 / ピクセルサイズ(公称値): 1.34 Å / ピクセルサイズ(実測値): 1.34 Å 詳細: The local resolution for chains k and l (lower-case chain IDs) is worse than the rest of the map. Therefore, these chains were docked as a rigid body into an 8 Angstrom-resolution low-pass ...詳細: The local resolution for chains k and l (lower-case chain IDs) is worse than the rest of the map. Therefore, these chains were docked as a rigid body into an 8 Angstrom-resolution low-pass filtered map and then edited to remove clashes. The quality of this portion of the model is to be considered lower than the rest of the model. 対称性のタイプ: POINT | ||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT / 空間: RECIPROCAL / Target criteria: R-factor, FSC / 詳細: REFINEMENT PROTOCOL--flexible | ||||||||||||||||||||
| 原子モデル構築 | 3D fitting-ID: 1 / Source name: PDB / タイプ: experimental model
| ||||||||||||||||||||
| 精密化 | 詳細: HYDROGENS HAVE BEEN ADDED IN THEIR RIDING POSITIONS | ||||||||||||||||||||
| 精密化ステップ | サイクル: LAST
|


