Inhibition of replication initiation of damaged DNA by RB1/E2F1 / ERKs are inactivated / Initiation of Nuclear Envelope (NE) Reformation / Cyclin A/B1/B2 associated events during G2/M transition / Co-inhibition by CTLA4 / Co-stimulation by CD28 / Beta-catenin phosphorylation cascade / Disassembly of the destruction complex and recruitment of AXIN to the membrane / Negative regulation of MAPK pathway / Spry regulation of FGF signaling ...Inhibition of replication initiation of damaged DNA by RB1/E2F1 / ERKs are inactivated / Initiation of Nuclear Envelope (NE) Reformation / Cyclin A/B1/B2 associated events during G2/M transition / Co-inhibition by CTLA4 / Co-stimulation by CD28 / Beta-catenin phosphorylation cascade / Disassembly of the destruction complex and recruitment of AXIN to the membrane / Negative regulation of MAPK pathway / Spry regulation of FGF signaling / Regulation of TP53 Degradation / RAF activation / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / meiotic spindle elongation / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / PP2A-mediated dephosphorylation of key metabolic factors / RNA polymerase II CTD heptapeptide repeat S2 phosphatase activity / RNA polymerase II CTD heptapeptide repeat S7 phosphatase activity / peptidyl-threonine dephosphorylation / Cyclin D associated events in G1 / DARPP-32 events / Turbulent (oscillatory, disturbed) flow shear stress activates signaling by PIEZO1 and integrins in endothelial cells / PKR-mediated signaling / regulation of meiotic cell cycle process involved in oocyte maturation / mitotic sister chromatid separation / MASTL Facilitates Mitotic Progression / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / protein phosphatase type 2A complex / Resolution of Sister Chromatid Cohesion / Degradation of beta-catenin by the destruction complex / meiotic sister chromatid cohesion, centromeric / INTAC complex / Platelet sensitization by LDL / ERK/MAPK targets / RNA polymerase II CTD heptapeptide repeat S5 phosphatase activity / RHO GTPases Activate Formins / Loss of Nlp from mitotic centrosomes / Recruitment of mitotic centrosome proteins and complexes / Loss of proteins required for interphase microtubule organization from the centrosome / FAR/SIN/STRIPAK complex / PI5P, PP2A and IER3 Regulate PI3K/AKT Signaling / Anchoring of the basal body to the plasma membrane / Separation of Sister Chromatids / Recruitment of NuMA to mitotic centrosomes / AURKA Activation by TPX2 / female meiotic nuclear division / Regulation of glycolysis by fructose 2,6-bisphosphate metabolism / meiotic sister chromatid cohesion / Inhibition of replication initiation of damaged DNA by RB1/E2F1 / Regulation of PLK1 Activity at G2/M Transition / protein phosphatase regulator activity / protein antigen binding / GABA receptor binding / APC truncation mutants have impaired AXIN binding / AXIN missense mutants destabilize the destruction complex / Truncations of AMER1 destabilize the destruction complex / positive regulation of extrinsic apoptotic signaling pathway in absence of ligand / ERKs are inactivated / Initiation of Nuclear Envelope (NE) Reformation / Beta-catenin phosphorylation cascade / Signaling by GSK3beta mutants / CTNNB1 S33 mutants aren't phosphorylated / CTNNB1 S37 mutants aren't phosphorylated / CTNNB1 S45 mutants aren't phosphorylated / CTNNB1 T41 mutants aren't phosphorylated / Co-stimulation by CD28 / RNA polymerase II transcription initiation surveillance / regulation of growth / Disassembly of the destruction complex and recruitment of AXIN to the membrane / protein dephosphorylation / negative regulation of epithelial to mesenchymal transition / Co-inhibition by CTLA4 / Platelet sensitization by LDL / protein-serine/threonine phosphatase / negative regulation of glycolytic process through fructose-6-phosphate / ERK/MAPK targets / vascular endothelial cell response to oscillatory fluid shear stress / protein serine/threonine phosphatase activity / T cell homeostasis / mesoderm development / positive regulation of NLRP3 inflammasome complex assembly / regulation of cell differentiation / regulation of microtubule polymerization / regulation of G1/S transition of mitotic cell cycle / protein phosphatase activator activity / lateral plasma membrane / intrinsic apoptotic signaling pathway in response to DNA damage by p53 class mediator / chromosome, centromeric region / DARPP-32 events / negative regulation of hippo signaling / Cyclin A/B1/B2 associated events during G2/M transition / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / spindle assembly / phosphoprotein phosphatase activity / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / protein tyrosine phosphatase activity / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / Turbulent (oscillatory, disturbed) flow shear stress activates signaling by PIEZO1 and integrins in endothelial cells Similarity search - Function
The biological assembly is a heterotrimer from one A subunit, one B subunit, and one C subunit. There are two heterotrimers in one asymmetric unit. each heterotrimer has one microcystin-LR, PP2A inhibitor.
-
Components
#1: Protein
Serine/threonine-proteinphosphatase2A65kDaregulatorysubunitAalphaisoform / PP2A / subunit A / PR65-alpha isoform / PP2A / subunit A / R1-alpha isoform
Mass: 65392.367 Da / Num. of mol.: 2 / Fragment: Aalpha subunit Source method: isolated from a genetically manipulated source Source: (gene. exp.) Mus musculus (house mouse) / Gene: Ppp2r1a / Plasmid: pGEX4T1 / Production host: Escherichia coli (E. coli) / Strain (production host): BL21 star References: UniProt: Q76MZ3, protein-serine/threonine phosphatase
#2: Protein
Serine/threonine-proteinphosphatase2A56kDaregulatorysubunitgammaisoform / PP2A / B subunit / B' gamma isoform / PP2A / B subunit / B56 gamma isoform / PP2A / B subunit / ...PP2A / B subunit / B' gamma isoform / PP2A / B subunit / B56 gamma isoform / PP2A / B subunit / PR61 gamma isoform / PP2A / B subunit / R5 gamma isoform / NY-REN-29 antigen
Mass: 48186.934 Da / Num. of mol.: 2 / Fragment: B56gamma1 subunit, residues 30-436 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PPP2R5C, KIAA0044 / Plasmid: pGEX4T1 / Production host: Escherichia coli (E. coli) / Strain (production host): BL21 star References: UniProt: Q13362, protein-serine/threonine phosphatase
#3: Protein
Serine/threonine-proteinphosphatase2Acatalyticsubunitalphaisoform / PP2A-alpha / Replication protein C / RP-C
Mass: 35649.195 Da / Num. of mol.: 2 / Fragment: Calpha subunit / Mutation: D88N Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PPP2CA / Production host: Trichoplusia ni (cabbage looper) References: UniProt: P67775, protein-serine/threonine phosphatase
Monochromator: Double-crystal, Si(111) / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray
Radiation wavelength
Wavelength: 0.9793 Å / Relative weight: 1
Reflection
Redundancy: 8.5 % / Av σ(I) over netI: 5.9 / Number: 554144 / Rmerge(I) obs: 0.161 / Χ2: 1.4 / D res high: 3.7 Å / D res low: 50 Å / Num. obs: 65390 / % possible obs: 98.8
Diffraction reflection shell
Highest resolution (Å)
Lowest resolution (Å)
% possible obs (%)
ID
Rmerge(I) obs
Chi squared
Redundancy
7.96
50
96
1
0.053
3.077
8.5
6.32
7.96
97.7
1
0.117
1.851
8.6
5.53
6.32
98.4
1
0.187
1.303
8.7
5.02
5.53
98.7
1
0.212
1.204
8.7
4.66
5.02
99.1
1
0.217
1.168
8.7
4.39
4.66
99.3
1
0.256
1.134
8.7
4.17
4.39
99.5
1
0.333
1.092
8.7
3.99
4.17
99.6
1
0.495
1.046
8.7
3.83
3.99
99.8
1
0.68
1.015
8.5
3.7
3.83
99.6
1
0.916
1.001
7.1
Reflection
Resolution: 3.5→50 Å / Num. obs: 76332 / % possible obs: 97.5 % / Redundancy: 4.4 % / Rmerge(I) obs: 0.118 / Net I/σ(I): 7.5
Reflection shell
Resolution (Å)
Redundancy (%)
Rmerge(I) obs
Diffraction-ID
% possible all
3.5-3.63
4.1
0.905
1
98.1
3.63-3.77
4.3
0.614
1
98.5
3.77-3.94
4.4
0.438
1
98.9
3.94-4.15
4.4
0.297
1
98.9
4.15-4.41
4.4
0.185
1
98.7
4.41-4.75
4.4
0.137
1
97.9
4.75-5.23
4.4
0.118
1
98
5.23-5.98
4.4
0.115
1
96.8
5.98-7.53
4.4
0.094
1
96.1
7.53-50
4.4
0.046
1
93
-
Phasing
Phasing
Method: SAD
Phasing dm
Method: Solvent flattening and Histogram matching / Reflection: 76221
Phasing dm shell
Resolution (Å)
Delta phi final
FOM
Reflection
18.19-100
78.2
0.643
518
13.13-18.19
66.2
0.816
886
10.8-13.13
65.4
0.858
1140
9.38-10.8
66.1
0.881
1318
8.41-9.38
65
0.892
1529
7.69-8.41
63.3
0.875
1679
7.13-7.69
65.3
0.855
1835
6.67-7.13
67.3
0.85
1946
6.29-6.67
67.2
0.843
2076
5.97-6.29
68.4
0.852
2227
5.7-5.97
71.7
0.854
2311
5.46-5.7
72.2
0.861
2456
5.24-5.46
71.5
0.869
2529
5.05-5.24
75
0.876
2674
4.88-5.05
76.2
0.883
2731
4.73-4.88
77.1
0.888
2871
4.59-4.73
79.7
0.901
2937
4.46-4.59
86.1
0.894
3007
4.34-4.46
90.7
0.896
3108
4.23-4.34
91.2
0.902
3203
4.13-4.23
90.4
0.902
3288
4.04-4.13
89.6
0.901
3294
3.95-4.04
91.2
0.901
3486
3.87-3.95
88.7
0.904
3516
3.79-3.87
90
0.907
3599
3.71-3.79
89.2
0.907
3603
3.65-3.71
88.7
0.908
3751
3.58-3.65
89.7
0.892
3761
3.5-3.58
90.6
0.775
4942
-
Processing
Software
Name
Version
Classification
NB
DENZO
datareduction
SCALEPACK
datascaling
SHARP
phasing
DM
6
phasing
REFMAC
refinement
PDB_EXTRACT
2
dataextraction
HKL-2000
datacollection
HKL-2000
datareduction
HKL-2000
datascaling
Refinement
Method to determine structure: SAD / Resolution: 3.5→20 Å / Cor.coef. Fo:Fc: 0.897 / Cor.coef. Fo:Fc free: 0.854 / SU B: 59.364 / SU ML: 0.455 / TLS residual ADP flag: LIKELY RESIDUAL / Cross valid method: THROUGHOUT / ESU R Free: 0.597 / Stereochemistry target values: MAXIMUM LIKELIHOOD / Details: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
Rfactor
Num. reflection
% reflection
Selection details
Rfree
0.31552
3832
5.1 %
RANDOM
Rwork
0.25681
-
-
-
obs
0.2598
72008
97.48 %
-
Solvent computation
Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.4 Å / Solvent model: MASK
Displacement parameters
Biso mean: 108.538 Å2
Refinement step
Cycle: LAST / Resolution: 3.5→20 Å
Protein
Nucleic acid
Ligand
Solvent
Total
Num. atoms
19951
0
4
0
19955
Refine LS restraints
Refine-ID
Type
Dev ideal
Dev ideal target
Number
X-RAY DIFFRACTION
r_bond_refined_d
0.009
0.022
20389
X-RAY DIFFRACTION
r_bond_other_d
X-RAY DIFFRACTION
r_angle_refined_deg
1.38
1.971
27690
X-RAY DIFFRACTION
r_angle_other_deg
X-RAY DIFFRACTION
r_dihedral_angle_1_deg
7.269
5
2511
X-RAY DIFFRACTION
r_dihedral_angle_2_deg
41.351
24.376
914
X-RAY DIFFRACTION
r_dihedral_angle_3_deg
22.445
15
3435
X-RAY DIFFRACTION
r_dihedral_angle_4_deg
16.513
15
114
X-RAY DIFFRACTION
r_chiral_restr
0.1
0.2
3190
X-RAY DIFFRACTION
r_gen_planes_refined
0.003
0.02
15326
X-RAY DIFFRACTION
r_gen_planes_other
X-RAY DIFFRACTION
r_nbd_refined
0.235
0.2
11543
X-RAY DIFFRACTION
r_nbd_other
X-RAY DIFFRACTION
r_nbtor_refined
0.312
0.2
14144
X-RAY DIFFRACTION
r_nbtor_other
X-RAY DIFFRACTION
r_xyhbond_nbd_refined
0.155
0.2
719
X-RAY DIFFRACTION
r_xyhbond_nbd_other
X-RAY DIFFRACTION
r_metal_ion_refined
0.032
0.2
1
X-RAY DIFFRACTION
r_metal_ion_other
X-RAY DIFFRACTION
r_symmetry_vdw_refined
0.271
0.2
43
X-RAY DIFFRACTION
r_symmetry_vdw_other
X-RAY DIFFRACTION
r_symmetry_hbond_refined
0.227
0.2
4
X-RAY DIFFRACTION
r_symmetry_hbond_other
X-RAY DIFFRACTION
r_symmetry_metal_ion_refined
X-RAY DIFFRACTION
r_symmetry_metal_ion_other
X-RAY DIFFRACTION
r_mcbond_it
0.363
1.5
12916
X-RAY DIFFRACTION
r_mcbond_other
X-RAY DIFFRACTION
r_mcangle_it
0.671
2
20428
X-RAY DIFFRACTION
r_scbond_it
0.765
3
8308
X-RAY DIFFRACTION
r_scangle_it
1.356
4.5
7262
X-RAY DIFFRACTION
r_rigid_bond_restr
X-RAY DIFFRACTION
r_sphericity_free
X-RAY DIFFRACTION
r_sphericity_bonded
LS refinement shell
Resolution: 3.5→3.588 Å / Total num. of bins used: 20
Rfactor
Num. reflection
% reflection
Rfree
0.332
253
-
Rwork
0.278
5195
-
obs
-
-
97.92 %
Refinement TLS params.
Method: refined / Refine-ID: X-RAY DIFFRACTION
ID
L11 (°2)
L12 (°2)
L13 (°2)
L22 (°2)
L23 (°2)
L33 (°2)
S11 (Å °)
S12 (Å °)
S13 (Å °)
S21 (Å °)
S22 (Å °)
S23 (Å °)
S31 (Å °)
S32 (Å °)
S33 (Å °)
T11 (Å2)
T12 (Å2)
T13 (Å2)
T22 (Å2)
T23 (Å2)
T33 (Å2)
Origin x (Å)
Origin y (Å)
Origin z (Å)
1
1.7024
-0.7702
-0.061
1.4094
-0.1957
0.2101
0.0732
0.4694
-0.4192
-0.6148
-0.1786
0.5373
0.0092
-0.1773
0.1054
0.0529
0.0904
-0.2281
0.1069
-0.269
-0.3051
118.0312
220.9926
98.3446
2
0.89
-0.1591
-0.3709
1.2606
0.4458
1.8055
-0.2379
-0.1023
-0.5066
0.1225
0.0041
0.2632
0.0921
-0.3855
0.2338
-0.5581
-0.122
0.2423
-0.6365
-0.005
-0.2814
166.6063
168.5954
107.2622
3
3.7098
-0.0788
-0.2996
1.7602
-0.2402
3.266
-0.0745
0.333
0.054
-0.3204
0.0391
0.038
0.0248
-0.1442
0.0354
-0.0667
0.2689
-0.2171
-0.1745
-0.1242
-0.1876
114.428
249.235
116.203
4
5.4633
0.6814
-0.0719
4.1511
-0.5067
6.0579
-0.0386
-0.1262
-0.4762
0.3322
-0.108
0.0171
0.4233
0.0498
0.1467
-1.0146
-0.0477
0.2552
-1.5077
-0.3204
-1.2523
194.8149
165.2957
86.1918
5
2.9339
-1.8352
-0.1022
3.4823
0.2217
0.8555
0.1333
-0.0602
0.2818
0.0602
-0.0688
-0.3129
-0.0134
0.2509
-0.0644
0.0643
-0.0644
0.1324
0.0599
-0.0421
-0.1589
195.4888
194.3911
116.4937
6
1.4718
0.1804
-0.2016
5.4423
-1.6328
2.0073
-0.177
-0.1561
0.2922
-0.1848
0.1289
-0.2331
-0.1295
-0.1522
0.0481
-0.0508
0.1295
0.0906
-0.2863
-0.0521
-0.2816
148.4674
227.0913
122.9527
Refinement TLS group
ID
Refine-ID
Refine TLS-ID
Auth asym-ID
Label asym-ID
Auth seq-ID
Label seq-ID
1
X-RAY DIFFRACTION
1
A
A
7 - 589
7 - 589
2
X-RAY DIFFRACTION
2
D
D
7 - 589
7 - 589
3
X-RAY DIFFRACTION
3
C
C
4 - 309
4 - 309
4
X-RAY DIFFRACTION
4
F
F
4 - 294
4 - 294
5
X-RAY DIFFRACTION
5
E
E
30 - 406
1 - 377
6
X-RAY DIFFRACTION
6
B
B
31 - 406
2 - 377
+
About Yorodumi
-
News
-
Feb 9, 2022. New format data for meta-information of EMDB entries
New format data for meta-information of EMDB entries
Version 3 of the EMDB header file is now the official format.
The previous official version 1.9 will be removed from the archive.
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information
 Homo sapiens (human)
Homo sapiens (human) Microcystis aeruginosa (bacteria)
Microcystis aeruginosa (bacteria) X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads





 PDBj
PDBj














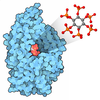









 Assembly
Assembly


 Trichoplusia ni (cabbage looper)
Trichoplusia ni (cabbage looper)
 Type: Oligopeptide / Class: Toxin / Mass: 1014.195 Da / Num. of mol.: 2 / Source method: obtained synthetically / Source: (synth.)
Type: Oligopeptide / Class: Toxin / Mass: 1014.195 Da / Num. of mol.: 2 / Source method: obtained synthetically / Source: (synth.) 
 Mass: 54.938 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mn
Mass: 54.938 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mn Sample preparation
Sample preparation / Beamline: 5.0.2 / Wavelength: 0.9793
/ Beamline: 5.0.2 / Wavelength: 0.9793  Processing
Processing