#1: Journal: Cell(Cambridge,Mass.) / Year: 2001 Title: The fusion glycoprotein shell of Semliki Forest Virus: an icosahedral assembly primed for fusogenic activation at endosomal pH.
#2: Journal: Nature / Year: 2004 Title: Conformational change and protein-protein interactions of the fusion protein of Semliki Forest virus.
History
Deposition
Aug 5, 2005
Deposition site: RCSB / Processing site: RCSB
Revision 1.0
Jan 17, 2006
Provider: repository / Type: Initial release
Revision 1.1
Apr 30, 2008
Group: Version format compliance
Revision 1.2
Jul 13, 2011
Group: Advisory / Refinement description / Version format compliance
Resolution: 3→3.21 Å / Redundancy: 5.5 % / Rmerge(I) obs: 0.124 / Mean I/σ(I) obs: 5.4 / % possible all: 76.1
Reflection
*PLUS
Highest resolution: 3 Å / Lowest resolution: 39 Å / Num. measured all: 82854
Reflection shell
*PLUS
Highest resolution: 3 Å / % possible obs: 76.1 %
-
Processing
Software
Name
Version
Classification
REFMAC
5.2.0005
refinement
MOSFLM
datareduction
SCALEPACK
datascaling
MLPHARE
phasing
Refinement
Method to determine structure: MIR / Resolution: 3→39 Å / Cor.coef. Fo:Fc: 0.873 / Cor.coef. Fo:Fc free: 0.809 / SU B: 52.882 / SU ML: 0.494 / TLS residual ADP flag: LIKELY RESIDUAL / Cross valid method: THROUGHOUT / ESU R Free: 0.553 / Stereochemistry target values: MAXIMUM LIKELIHOOD Details: 8 OF THE SOLVENT MOLECULES ADDED TO POSITIVE FOURIER DIFFERENCE PEAKS (ABOVE 3 SIGMA) ARE FURTHER THAN 3.5 ANGSTROMS AWAY FROM MACROMOLECULE. THE E1 PROTEIN IS GLYCOSYLATED at ASN 141. THE ...Details: 8 OF THE SOLVENT MOLECULES ADDED TO POSITIVE FOURIER DIFFERENCE PEAKS (ABOVE 3 SIGMA) ARE FURTHER THAN 3.5 ANGSTROMS AWAY FROM MACROMOLECULE. THE E1 PROTEIN IS GLYCOSYLATED at ASN 141. THE DENSITY WAS NOT CLEAR ENOUGH TO BUILD A SUGAR AT THIS LOCATION THUS WATER MOLECULES 71 AND 81 WERE PLACED INTO POSITIVE DENSITY NEAR THIS RESIDUE.
Rfactor
Num. reflection
% reflection
Selection details
Rfree
0.31868
1232
10.4 %
RANDOM
Rwork
0.26654
-
-
-
obs
0.27199
10604
87.9 %
-
all
-
13467
-
-
Solvent computation
Ion probe radii: 0.8 Å / Shrinkage radii: 0.8 Å / VDW probe radii: 1.2 Å / Solvent model: MASK
Displacement parameters
Biso mean: 4.062 Å2
Baniso -1
Baniso -2
Baniso -3
1-
-3.51 Å2
-1.76 Å2
0 Å2
2-
-
-3.51 Å2
0 Å2
3-
-
-
5.27 Å2
Refinement step
Cycle: LAST / Resolution: 3→39 Å
Protein
Nucleic acid
Ligand
Solvent
Total
Num. atoms
2935
0
0
88
3023
Refine LS restraints
Refine-ID
Type
Dev ideal
Dev ideal target
Number
X-RAY DIFFRACTION
r_bond_refined_d
0.006
0.022
3020
X-RAY DIFFRACTION
r_bond_other_d
0.001
0.02
2610
X-RAY DIFFRACTION
r_angle_refined_deg
0.996
1.947
4125
X-RAY DIFFRACTION
r_angle_other_deg
0.707
3
6121
X-RAY DIFFRACTION
r_dihedral_angle_1_deg
6.875
5
383
X-RAY DIFFRACTION
r_dihedral_angle_2_deg
36.652
24.298
121
X-RAY DIFFRACTION
r_dihedral_angle_3_deg
15.366
15
466
X-RAY DIFFRACTION
r_dihedral_angle_4_deg
10.636
15
10
X-RAY DIFFRACTION
r_chiral_restr
0.06
0.2
464
X-RAY DIFFRACTION
r_gen_planes_refined
0.002
0.02
3366
X-RAY DIFFRACTION
r_gen_planes_other
0.001
0.02
589
X-RAY DIFFRACTION
r_nbd_refined
0.175
0.2
697
X-RAY DIFFRACTION
r_nbd_other
0.158
0.2
2872
X-RAY DIFFRACTION
r_nbtor_refined
0.173
0.2
1471
X-RAY DIFFRACTION
r_nbtor_other
0.078
0.2
1858
X-RAY DIFFRACTION
r_xyhbond_nbd_refined
0.153
0.2
113
X-RAY DIFFRACTION
r_xyhbond_nbd_other
0.058
0.2
3
X-RAY DIFFRACTION
r_metal_ion_refined
X-RAY DIFFRACTION
r_metal_ion_other
X-RAY DIFFRACTION
r_symmetry_vdw_refined
0.124
0.2
14
X-RAY DIFFRACTION
r_symmetry_vdw_other
0.101
0.2
43
X-RAY DIFFRACTION
r_symmetry_hbond_refined
0.201
0.2
7
X-RAY DIFFRACTION
r_symmetry_hbond_other
X-RAY DIFFRACTION
r_symmetry_metal_ion_refined
X-RAY DIFFRACTION
r_symmetry_metal_ion_other
X-RAY DIFFRACTION
r_mcbond_it
0.184
1.5
2457
X-RAY DIFFRACTION
r_mcbond_other
0.017
1.5
768
X-RAY DIFFRACTION
r_mcangle_it
0.21
2
3129
X-RAY DIFFRACTION
r_scbond_it
0.246
3
1310
X-RAY DIFFRACTION
r_scangle_it
0.378
4.5
996
X-RAY DIFFRACTION
r_rigid_bond_restr
X-RAY DIFFRACTION
r_sphericity_free
X-RAY DIFFRACTION
r_sphericity_bonded
LS refinement shell
Resolution: 3→3.077 Å / Total num. of bins used: 20
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information
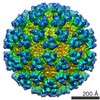 Semliki forest virus
Semliki forest virus X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads










 PDBj
PDBj
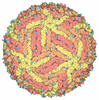

 Assembly
Assembly
 Mass: 18.015 Da / Num. of mol.: 88 / Source method: isolated from a natural source / Formula: H2O
Mass: 18.015 Da / Num. of mol.: 88 / Source method: isolated from a natural source / Formula: H2O Sample preparation
Sample preparation / Beamline: BM14 / Wavelength: 0.945 Å
/ Beamline: BM14 / Wavelength: 0.945 Å Processing
Processing