 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 1qzv | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Crystal structure of plant photosystem I | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | PHOTOSYNTHESIS / PLANT PHOTOSYNTHETIC REACTION CENTER / PERIPHERAL ANTENNA / LIGHT-HARVESTING SYSTEM / PLANT MEMBRANE PROTEIN COMPLEX | |||||||||
| Function / homology | CHLOROPHYLL A / PHYLLOQUINONE / IRON/SULFUR CLUSTER Function and homology information Function and homology information | |||||||||
| Biological species |  Pisum sativum (garden pea) Pisum sativum (garden pea) | |||||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / MIR / Resolution: 4.44 Å X-RAY DIFFRACTION / SYNCHROTRON / MIR / Resolution: 4.44 Å | |||||||||
 Authors Authors | Ben-Shem, A. / Frolow, F. / Nelson, N. | |||||||||
 Citation Citation | Journal: Nature / Year: 2003 Title: Crystal structure of plant photosystem I. Authors: Ben-Shem, A. / Frolow, F. / Nelson, N. | |||||||||
| History |
| |||||||||
| Remark 999 | SEQUENCE AND NOMENCLATURE: THE NUMBERING OF RESIDUES IN CHAINS A,B,C,D,E,F,I,J,K,L IS DERIVED FROM ... SEQUENCE AND NOMENCLATURE: THE NUMBERING OF RESIDUES IN CHAINS A,B,C,D,E,F,I,J,K,L IS DERIVED FROM PAIR-WISE ALIGNMENT BETWEEN THE SEQUENCE IN HIGHER PLANTS AND THAT IN CYANOBACTERIA (SYNECHOCOCCUS ELONGATUS) AND FROM COMPARISON OF THE PLANT STRUCTURE TO THE CYANOBACTERIAL STRUCTURE (PDB CODE 1JB0). SINCE THE RESULTS FROM SUCH A PROCEDURE MIGHT NOT BE EXACT, RESIDUE TYPE IN MOST CASES IS DESIGNATED AS UNKNOWN (UNK). IN CHAIN D RESIDUES D1-D22 CORRESPOND TO AN ADDITIONAL N-TERMINAL DOMAIN THAT HAS NO PARALLEL IN CYANOBACTERIA. DUE TO DISCONTINUITY IN THE MAP AND THE MODEL (NEXT RESIDUE IS D31) THE POSSIBILITY EXISTS THAT THE DIRECTION OF THIS SEGMENT IS ACTUALLY D22 -> D1. NUMBERING OF RESIDUES IN CHAINS H AND G SHOULD BE REGARDED AS SPECULATIVE. MOST SECONDARY STRUCTURE PREDICTING PROGRAMS SUGGEST THAT THE TRANSMEMBRANE HELIX OF CHAIN H IS LOCATED TOWARDS THE C-TERMINUS OF THIS CHAIN. THIS SUGGESTION WAS ADOPTED HERE. SUBUNIT G HAS TWO TRANSMEMBRANe HELICES - ONE TOWARDS THE N-TERMINUS AND THE OTHER TOWARDS THE C-TERMINUS. OUR CHOICE REGARDING WHICH OF THE TWO OBSERVED HELICES SHOULD BE ASSIGNED TO THE N-TERMINUS AND WHICH TO THE C-TERMINUS IS ARBITRARY. NUMBERING OF RESIDUES IN THE FOUR LHCI SUBUNITS IS BASED ON MULTIPLE SEQUENCE ALIGNMENT OF SEVERAL LHCI AND LHCII PROTEINS (SEE MELKOZERNOV AND BLANKENSHIP 2003). WE IDENTIFIED IN THE MODEL AND IN THE ALIGNMENT THE RESIDUES THAT SEEM TO BE THE LIGANDS OF CHLOROPHYLLS A1,A2,A3,A4,A5,A6,B3,B5,B6 (LABELING AS IN LHCII). THESE ANCHOR POINTS DETERMINED RESIDUE NUMBERING OF HELICES A,B,C,D AND THEIR CONTINUATIONS IN ALL LHCI SUBUNITS. AS FOR RESIDUE NUMBERS IN LOOPS THAT ARE NOT CONNECTED TO THE HELICES IN THE MODEL (DISCONTINUITIES)- THESE ARE SPECULATIVE. IN LHCA3 ONLY, THE TRANSMEMBRANE HELIX CLOSEST TO HELIX C IS NOT THE LONGEST OF THE TWO TILTED TRANSMEMBRANE HELICES (LABELING AS IN KUHLBRANDT ET AL. 1994). THIS MIGHT INDICATE THAT IN LHCA3 THE HELIX CLOSEST TO HELIX C IS NOT THE FIRST HELIX (FROM THE N-TERMINUS) BUT THE THIRD ONE. THE ELECTRON DENSITY MAP IS NOT CONCLUSIVE HERE BUT SEEMS TO SUGGEST THAT THIS MIGHT BE THE CASE. IF THIS IS TRUE, AND LHCA3 FOLDS DIFFERENTLY, THEN THE NUMBERING OF RESIDUES IN LHCA3 MUST CHANGE ACCORDINGLY. C-ALPHA ATOMS ONLY WERE SUBMITTED FOR THIS MODEL. CHLOROPHYLL NOTATION: REACTION CENTER CHLOROPHYLLS - FOLLOW THE NOTATION OF THE CYANOBACTERIAL STRUCTURE PDB CODE 1JB0 (NO CHAIN I.D.) "GAP" CHLOROPHYLLS (SEE BEN-SHEM ET AL. NATURE 426 630-635) - NO CHAIN I.D., RESIDUE NUMBERS 4001-4010. THESE DO NOT INCLUDE "LINKER" CHLOROPHYLLS. LHCI CHLOROPHYLLS - CHAIN I.D. 1,2,3,4 CORRESPONDING TO LHCA1-4. RESIDUE NUMBERS 1011-1017 IDENTIFY CHLOROPHYLLS AT POSITIONS SIMILAR TO A1-A7 IN LHCII. RESIDUE NUMBERS 1021-1026 IDENTIFY CHLOROPHYLLS AT POSITIONS SIMILAR TO B1-B6 IN LHCII, RESIDUE NUMBERS 1031-1033 IDENTIFY "LINKER" CHLOROPHYLLS. RESIDUE NUMBER 1041 IDENTIFY ONE CHLOROPHYLL IN LHCA3 WHICH IS IN A POSITION BETWEEN A3 AND B3. CHLOROPHYLLS WITH CHAIN I.D. 7,8,9,0 ARE THE SYMMETRY EQUIVALENTS OF CHLOROPHYLLS WITH CHAIN I.D. 1,2,3,4. CHLOROPHYLLS NUMBERED 5011-5901 ARE THE SYMMETRY EQUIVALENTS OF CHLOROPHYLLS NUMBERED 1011-1901. CHLOROPHYLLS NUMBERED 8001-8010 ARE THE SYMMETRY EQUIVALENTS OF CHLOROPHYLLS NUMBERED 4001-4010. PQN 6001 AND 6002 ARE THE SYMMETRY EQUIVALENTS OF PQN 2001 AND 2002. FS4 MOLECULES 7001-7003 ARE THE SYMMETRY EQUIVALENTS OF FS4 MOLECULES 3001-3003. | |||||||||
| Remark 600 | HETEROGEN ATOMS C11, C12, C13, C14, C15, C16, C17, C18, C19, C20, C21, C22, C23, C24, C25, C26, ...HETEROGEN ATOMS C11, C12, C13, C14, C15, C16, C17, C18, C19, C20, C21, C22, C23, C24, C25, C26, C27, C28, C29, AND C30 FOR LIGAND PQN ARE MISSING IN THE COORDINATES. ATOMS CMA, CAA, CBA, CGA, O1A, O2A, CMB, CAB, CBB, CMC, CAC, CBC, CMD, CAD, OBD, CBD, CGD, O1D, O2D, CED, C1, C2, C3, C4, C5, C6, C7, C8, C9, C10, C11, C12, C13, C14, C15, C16, C17, C18, C19, AND C20 FOR LIGAND CLA ARE MISSING IN THE COORDINATES. CHLOROPHYLLS WERE MODELLED BY PORPHYRINS. | |||||||||
| Remark 400 | COMPOUND THE ASYMMETRIC UNIT CONTAINS 2 MONOMERS. EACH MONOMER IS A BIOLOGICAL UNIT. MONOMER ONE ... COMPOUND THE ASYMMETRIC UNIT CONTAINS 2 MONOMERS. EACH MONOMER IS A BIOLOGICAL UNIT. MONOMER ONE CONSISTS OF CHAINS A-L AND 1-4. MONOMER TWO CONSISTS OF CHAINS P-Z AND 5-0. CHAIN IDS P-Z AND 5-0 ARE FOR THE CHAIN ID ASSIGNMENT IN THE PDB FILE ONLY AND ARE NOT SUPPOSED TO IMPLY THEY ARE SUBUNITS THAT ARE NOT PART OF THE FIRST MONOMER. THE CHAINS ARE RELATED BY NCS IN THE FOLLOWING WAY: CHAIN P IS THE NCS PARTNER OF CHAIN A. CHAIN Q IS THE NCS PARTNER OF CHAIN B. CHAIN R IS THE NCS PARTNER OF CHAIN C. CHAIN S IS THE NCS PARTNER OF CHAIN D. CHAIN T IS THE NCS PARTNER OF CHAIN E. CHAIN U IS THE NCS PARTNER OF CHAIN F. CHAIN V IS THE NCS PARTNER OF CHAIN G. CHAIN W IS THE NCS PARTNER OF CHAIN H. CHAIN Y IS THE NCS PARTNER OF CHAIN I. CHAIN Z IS THE NCS PARTNER OF CHAIN J. CHAIN 5 IS THE NCS PARTNER OF CHAIN K. CHAIN 6 IS THE NCS PARTNER OF CHAIN L. CHAIN 7 IS THE NCS PARTNER OF CHAIN 1. CHAIN 8 IS THE NCS PARTNER OF CHAIN 2. CHAIN 9 IS THE NCS PARTNER OF CHAIN 3. CHAIN 0 IS THE NCS PARTNER OF CHAIN 4. | |||||||||
| Remark 295 | NON-CRYSTALLOGRAPHIC SYMMETRY THE TRANSFORMATIONS PRESENTED ON THE MTRIX RECORDS BELOW DESCRIBE ... NON-CRYSTALLOGRAPHIC SYMMETRY THE TRANSFORMATIONS PRESENTED ON THE MTRIX RECORDS BELOW DESCRIBE NON-CRYSTALLOGRAPHIC RELATIONSHIPS AMONG ATOMS IN THIS ENTRY. APPLYING THE APPROPRIATE MTRIX TRANSFORMATION TO THE RESIDUES LISTED FIRST WILL YIELD APPROXIMATE COORDINATES FOR THE RESIDUES LISTED SECOND. CHAIN IDENTIFIERS GIVEN AS "?" REFER TO CHAINS FOR WHICH ATOMS ARE NOT FOUND IN THIS ENTRY. APPLIED TO TRANSFORMED TO TRANSFORM CHAIN RESIDUES CHAIN RESIDUES RMSD SSS M 1 A 29 .. 758 P 29 .. 758 0.000 WHERE SSS -> COLUMNS 8-10 OF MTRIX RECORDS REMARK: THE RMSD IS 0.00 IN ALL TRANSFORMED CHAINS. THEY WERE ALL GENERATED AUTOMATICALLY BY EXACTLY THE SAME TRANSFORMATION, FROM THE FIRST MONOMER. |
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 1qzv.cif.gz | 454.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb1qzv.ent.gz | 324.5 KB | Display | PDB format |
| PDBx/mmJSON format | 1qzv.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/qz/1qzvftp://data.pdbj.org/pub/pdb/validation_reports/qz/1qzv | HTTPS FTP |
|---|
-Related structure data
| Similar structure data |
|---|




-Links
 PDBj
PDBj


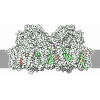
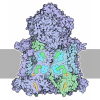
- Assembly
Assembly
| Deposited unit | 
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 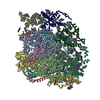
| ||||||||
| 2 | 
| ||||||||
| Unit cell |
| ||||||||
| Noncrystallographic symmetry (NCS) | NCS oper: (Code: given Matrix: (0.82712, -0.02534, 0.56146), Vector: |
-Components
-PLANT PHOTOSYSTEM I: SUBUNIT ... , 12 types, 24 molecules APBQCRDSETFUGVHWIYJZK5L6
| #1: Protein | Mass: 61804.074 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#2: Protein | Mass: 62314.688 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#3: Protein | Mass: 6826.406 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#4: Protein | Mass: 13124.170 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#5: Protein | Mass: 5464.728 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#6: Protein | Mass: 13804.815 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#7: Protein | Mass: 6315.777 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#8: Protein | Mass: 4443.468 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#9: Protein/peptide | Mass: 2571.161 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#10: Protein/peptide | Mass: 3507.314 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#11: Protein/peptide | Mass: 3592.419 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#12: Protein | Mass: 11507.176 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA |
|---|
-PLANT LIGHT HARVESTING COMPLEX I(LHCI): SUBUNIT ... , 4 types, 8 molecules 17283940
| #13: Protein | Mass: 9294.448 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#14: Protein | Mass: 9805.078 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#15: Protein | Mass: 9975.288 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA#16: Protein | Mass: 10145.497 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) Pisum sativum (garden pea) / Cellular location: Thylakoid Membrane / Strain: ALASKA |
|---|
-Non-polymers , 3 types, 344 molecules 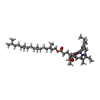
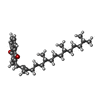
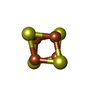
| #17: Chemical | ChemComp-CLA /  Mass: 893.489 Da / Num. of mol.: 334 / Source method: obtained synthetically / Formula: C55H72MgN4O5 Mass: 893.489 Da / Num. of mol.: 334 / Source method: obtained synthetically / Formula: C55H72MgN4O5#18: Chemical | ChemComp-PQN /  Mass: 450.696 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C31H46O2 Mass: 450.696 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: C31H46O2#19: Chemical | ChemComp-SF4 /  Mass: 351.640 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Fe4S4 Mass: 351.640 Da / Num. of mol.: 6 / Source method: obtained synthetically / Formula: Fe4S4 |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 3 |
|---|
- Sample preparation
Sample preparation
| Crystal grow | Temperature: 277 K / Method: vapor diffusion, sitting drop / pH: 6.6 Details: Ben-Shem, A., Nelson, N., Frolow, F. (2003) Acta Crystallogr D. 59:1824-7. PEG 6000, AMMONIUM CITRATE, PEG 400, MES, BIS-TRIS, DODECYL-THIO-MALTOSIDE, pH 6.6, VAPOR DIFFUSION, SITTING DROP, temperature 277K | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Crystal grow | *PLUS Temperature: 277 K / Method: vapor diffusion, sitting drop / Details: Ben-Shem, A., (2003) Acta Cryst., D59, 1824. | ||||||||||||||||||||||||||||||||||||
| Components of the solutions | *PLUS
|
-Data collection
| Diffraction |
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diffraction source |
| |||||||||||||||
| Detector |
| |||||||||||||||
| Radiation | Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray | |||||||||||||||
| Radiation wavelength |
| |||||||||||||||
| Reflection | Resolution: 4.44→50 Å / Num. all: 92318 / Num. obs: 92318 / % possible obs: 99.6 % / Observed criterion σ(F): 0 / Observed criterion σ(I): -3 / Redundancy: 9 % / Rmerge(I) obs: 0.099 / Rsym value: 0.099 / Net I/σ(I): 18.8 | |||||||||||||||
| Reflection shell | Resolution: 4.44→4.52 Å / Redundancy: 7 % / Rmerge(I) obs: 0.86 / Mean I/σ(I) obs: 2.05 / Num. unique all: 4562 / Rsym value: 0.86 / % possible all: 99.5 |

- Processing
Processing
| Software |
| |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: MIR / Resolution: 4.44→50 Å / σ(F): 0 / Stereochemistry target values: REFMAC MONOMER LIBRARY Details: RIGID BODY REFINEMENT, BULK SOLVENT CORRECTION AND TLS REFINEMENT (OF CORE REGION).
| |||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 4.44→50 Å
|
