Journal: J Virol / Year: 2019 Title: Medusavirus, a Novel Large DNA Virus Discovered from Hot Spring Water. Authors: Genki Yoshikawa / Romain Blanc-Mathieu / Chihong Song / Yoko Kayama / Tomohiro Mochizuki / Kazuyoshi Murata / Hiroyuki Ogata / Masaharu Takemura / Abstract: Recent discoveries of new large DNA viruses reveal high diversity in their morphologies, genetic repertoires, and replication strategies. Here, we report the novel features of medusavirus, a large ...Recent discoveries of new large DNA viruses reveal high diversity in their morphologies, genetic repertoires, and replication strategies. Here, we report the novel features of medusavirus, a large DNA virus newly isolated from hot spring water in Japan. Medusavirus, with a diameter of 260 nm, shows a T=277 icosahedral capsid with unique spherical-headed spikes on its surface. It has a 381-kb genome encoding 461 putative proteins, 86 of which have their closest homologs in , whereas 279 (61%) are orphan genes. The virus lacks the genes encoding DNA topoisomerase II and RNA polymerase, showing that DNA replication takes place in the host nucleus, whereas the progeny virions are assembled in the cytoplasm. Furthermore, the medusavirus genome harbored genes for all five types of histones (H1, H2A, H2B, H3, and H4) and one DNA polymerase, which are phylogenetically placed at the root of the eukaryotic clades. In contrast, the host amoeba encoded many medusavirus homologs, including the major capsid protein. These facts strongly suggested that amoebae are indeed the most promising natural hosts of medusavirus, and that lateral gene transfers have taken place repeatedly and bidirectionally between the virus and its host since the early stage of their coevolution. Medusavirus reflects the traces of direct evolutionary interactions between the virus and eukaryotic hosts, which may be caused by sharing the DNA replication compartment and by evolutionarily long lasting virus-host relationships. Based on its unique morphological characteristics and phylogenomic relationships with other known large DNA viruses, we propose that medusavirus represents a new family, We have isolated a new nucleocytoplasmic large DNA virus (NCLDV) from hot spring water in Japan, named medusavirus. This new NCLDV is phylogenetically placed at the root of the eukaryotic clades based on the phylogenies of several key genes, including that encoding DNA polymerase, and its genome surprisingly encodes the full set of histone homologs. Furthermore, its laboratory host, , encodes many medusavirus homologs in its genome, including the major capsid protein, suggesting that the amoeba is the genuine natural host from ancient times of this newly described virus and that lateral gene transfers have repeatedly occurred between the virus and amoeba. These results suggest that medusavirus is a unique NCLDV preserving ancient footprints of evolutionary interactions with its hosts, thus providing clues to elucidate the evolution of NCLDVs, eukaryotes, and virus-host interaction. Based on the dissimilarities with other known NCLDVs, we propose that medusavirus represents a new viral family, .
History
Deposition
Aug 15, 2018
-
Header (metadata) release
Feb 6, 2019
-
Map release
Feb 6, 2019
-
Update
Apr 24, 2019
-
Current status
Apr 24, 2019
Processing site: PDBj / Status: Released
-
Structure visualization
Movie
Surface view with section colored by density value
Cryogen name: ETHANE / Chamber humidity: 95 % / Chamber temperature: 277 K / Instrument: FEI VITROBOT MARK IV
-
Electron microscopy
Microscope
JEOL 2200FS
Temperature
Min: 76.0 K / Max: 77.0 K
Specialist optics
Energy filter - Name: In-column Omega Filter / Energy filter - Slit width: 20 eV
Image recording
Film or detector model: DIRECT ELECTRON DE-20 (5k x 3k) / Digitization - Dimensions - Width: 5120 pixel / Digitization - Dimensions - Height: 3840 pixel / Digitization - Sampling interval: 6.4 µm / Digitization - Frames/image: 3-75 / Number real images: 1198 / Average exposure time: 3.0 sec. / Average electron dose: 20.0 e/Å2
Electron beam
Acceleration voltage: 200 kV / Electron source: FIELD EMISSION GUN
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Map data
Map data Sample
Sample Medusavirus
Medusavirus Authors
Authors Citation
Citation
 Structure visualization
Structure visualization Movie viewer
Movie viewer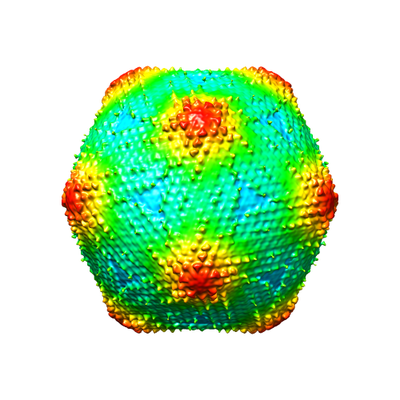
 Downloads & links
Downloads & links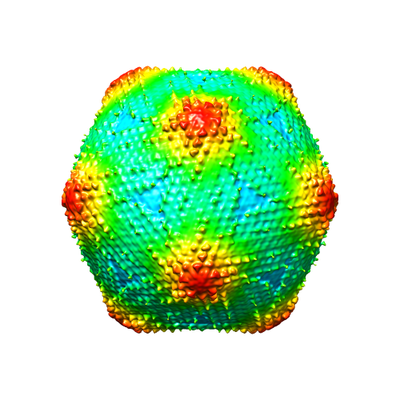 emd_9619.png
emd_9619.png http://ftp.pdbj.org/pub/emdb/structures/EMD-9619
http://ftp.pdbj.org/pub/emdb/structures/EMD-9619



 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)

















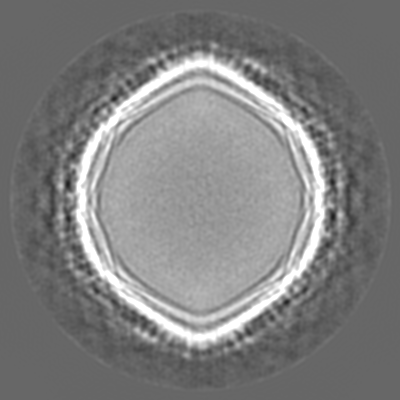


 Sample components
Sample components Acanthamoeba castellanii (eukaryote)
Acanthamoeba castellanii (eukaryote) Processing
Processing Electron microscopy
Electron microscopy FIELD EMISSION GUN
FIELD EMISSION GUN