 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Augmin/V junction(open) | |||||||||
 Map data Map data | ||||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | Augmin complex / PLANT PROTEIN | |||||||||
| Function / homology |  Function and homology information Function and homology informationmegagametogenesis / microgametogenesis / phragmoplast microtubule organization / cytokinesis by cell plate formation / HAUS complex / microtubule minus-end binding / phragmoplast / nuclear microtubule / microtubule organizing center organization / mitotic spindle microtubule ...megagametogenesis / microgametogenesis / phragmoplast microtubule organization / cytokinesis by cell plate formation / HAUS complex / microtubule minus-end binding / phragmoplast / nuclear microtubule / microtubule organizing center organization / mitotic spindle microtubule / microtubule nucleation / microtubule organizing center / spindle assembly / spindle microtubule / microtubule cytoskeleton organization / spindle / mitotic spindle / mitotic cell cycle / microtubule binding / microtubule / cell division / nucleus / cytoplasm Similarity search - Function | |||||||||
| Biological species |  | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 8.6 Å | |||||||||
 Authors Authors | Ashaduzzaman M / Al-Bassam J / Taheri A | |||||||||
| Funding support |  United States, 1 items United States, 1 items
| |||||||||
 Citation Citation | Journal: bioRxiv / Year: 2025 Title: Cryo-EM structures of the Plant Augmin reveal its intertwined coiled-coil assembly, antiparallel dimerization and NEDD1 binding mechanisms. Authors: Md Ashaduzzaman / Aryan Taheri / Yuh-Ru Julie Lee / Yuqi Tang / Fei Guo / Stephen D Fried / Bo Liu / Jawdat Al-Bassam / Abstract: Microtubule (MT) branch nucleation is fundamental for building parallel MT networks in eukaryotic cells. In plants and metazoans, MT branch nucleation requires Augmin and NEDD1 proteins which bind ...Microtubule (MT) branch nucleation is fundamental for building parallel MT networks in eukaryotic cells. In plants and metazoans, MT branch nucleation requires Augmin and NEDD1 proteins which bind along MTs and then recruit and activate the gamma-tubulin ring complex (γ-TuRC). Augmin is a fork-shaped assembly composed of eight coiled-coil subunits, while NEDD1 is a WD40 β-propellor protein that bridges across MTs, Augmin, and γ-TuRC during MT branch nucleation. Here, we reconstitute hetero-tetrameric and hetero-octameric Arabidopsis thaliana Augmin assemblies, resolve their subunit interactions using crosslinking mass spectrometry and determine 3.7 to 7.3-Å cryo-EM structures for the V-junction and extended regions of Augmin. These structures allowed us to generate a complete de novo plant Augmin model that reveals the long-range multi coiled-coil interfaces that stabilize its 40-nm hetero-octameric fork-shaped organization. We discovered the dual calponin homology (CH) domain forming its MT binding site at the end of its V-junction undertake open and closed conformations. We determined a 12-Å dimeric Augmin cryo-EM structure revealing Augmin undergoes anti-parallel dimerization through two conserved surfaces along Augmin's extended region. We reconstituted the NEDD1 WD40 β-propellor with Augmin revealing it directly binds on top its V-junction and enhances Augmin dimerization. Our studies suggest that cooperativity between the Augmin dual CH domains and NEDD1 WD40 binding site may regulate Augmin V-junction dual binding to MT lattices. This unique V-shaped dual binding and organization anchors Augmins along MTs generating a platform to recruit γ-TuRC and activate branched MT nucleation. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_49224.map.gz | 27.1 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-49224-v30.xmlemd-49224.xml | 25.5 KB 25.5 KB | Display Display | EMDB header |
| FSC (resolution estimation) | emd_49224_fsc.xml | 7.5 KB | Display | FSC data file |
| Images |  emd_49224.png emd_49224.png | 29.1 KB | ||
| Filedesc metadata | emd-49224.cif.gz | 7.9 KB | ||
| Others | emd_49224_half_map_1.map.gzemd_49224_half_map_2.map.gz | 28.3 MB 28.3 MB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-49224ftp://ftp.pdbj.org/pub/emdb/structures/EMD-49224 http://ftp.pdbj.org/pub/emdb/structures/EMD-49224ftp://ftp.pdbj.org/pub/emdb/structures/EMD-49224 | HTTPS FTP |
-Related structure data
| Related structure data |  9nbaMC  9na8C 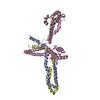 9na9C 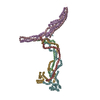 9nbbC 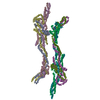 9nbdC M: atomic model generated by this map C: citing same article ( |
|---|---|
| Similar structure data |




-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|
-Map
| File | Download / File: emd_49224.map.gz / Format: CCP4 / Size: 30.5 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.76 Å | ||||||||||||||||||||||||||||||||||||
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-Supplemental data
-Half map: #2
| File | emd_49224_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
| Density Histograms |








-Half map: #1
| File | emd_49224_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
| Density Histograms |








- Sample components
Sample components
-Entire : AUGMIN, V-junction(open)
| Entire | Name: AUGMIN, V-junction(open) |
|---|---|
| Components |
|
-Supramolecule #1: AUGMIN, V-junction(open)
| Supramolecule | Name: AUGMIN, V-junction(open) / type: complex / ID: 1 / Parent: 0 / Macromolecule list: all |
|---|---|
| Source (natural) | Organism: |
| Molecular weight | Theoretical: 400 KDa |
-Macromolecule #1: AUGMIN subunit 2
| Macromolecule | Name: AUGMIN subunit 2 / type: protein_or_peptide / ID: 1 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: |
| Molecular weight | Theoretical: 33.057941 KDa |
| Recombinant expression | Organism:  |
| Sequence | String: MSMGGDTTWV GKKPIRRIGG LSDALSIASD LGFAVAPPPS QEELQSFASS NGERGDDLIR VLRELSVVQR KIADLQVELQ GRKDDKNVA HLTHVGEMQK KIETLSRITQ ILKDVIQNKD RIIARLQQPY SLDCIPVEAE YQKQFSELLM KAASDYGALT A SVSDFQWS ...String: MSMGGDTTWV GKKPIRRIGG LSDALSIASD LGFAVAPPPS QEELQSFASS NGERGDDLIR VLRELSVVQR KIADLQVELQ GRKDDKNVA HLTHVGEMQK KIETLSRITQ ILKDVIQNKD RIIARLQQPY SLDCIPVEAE YQKQFSELLM KAASDYGALT A SVSDFQWS QNFKEPPSVW GEMLRPIPVA LASCTRFFEA MSAMRESFAT LQELRVGNSA VSLPTTPGGN EMTHRDSDCV TP PQGRIES SFDDLAVHKT RRQNNDQNEE EEEEEEEEDG NNNGNRRLSW PPSVKKSSV UniProtKB: AUGMIN subunit 2 |
-Macromolecule #2: AUGMIN subunit 3
| Macromolecule | Name: AUGMIN subunit 3 / type: protein_or_peptide / ID: 2 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: |
| Molecular weight | Theoretical: 69.809453 KDa |
| Recombinant expression | Organism: |
| Sequence | String: MSSARLCSLV AELGYEGAGK LDPDSFEWPF QYDDARPILD WICSSLRPSN VLSLAELSLY EQFQRDGKLL EGDDLDQAYD SISAFSSRR NNQEAVFGAE ESIKEVRDAT LAHKAEALEL QRQLRRLQTQ YDLLTGQSSA LIQGRRARVA ATSAVSGQIT A IEDSLSAR ...String: MSSARLCSLV AELGYEGAGK LDPDSFEWPF QYDDARPILD WICSSLRPSN VLSLAELSLY EQFQRDGKLL EGDDLDQAYD SISAFSSRR NNQEAVFGAE ESIKEVRDAT LAHKAEALEL QRQLRRLQTQ YDLLTGQSSA LIQGRRARVA ATSAVSGQIT A IEDSLSAR NLQMNGVLGR LASTSQELAH YHSGEEDGIY LAYSDFHAYL AGDSACTKEL NQWFAKQLDT GPYRLVAEEG KS KCSWVSL DDTSNMLRDL EKSQHQRVAE LQRLRSIFGT SERQWIEAQV ENAKQQAILL TLKSQVTSVE AHIHFDLHSL RRK HADLVE EISTLYQKEE KLLSETIPEL CWELAQLQDT YILQGDYDLK VMRQELYISK QKVFINHLVN QLARHQFLKL ACQL EKKNM LGAFSLLKVI ESELQGYLSA TRSRVGRCSA LIQAASDVQE QGAVDDRDSF LHGVRDLLSI HSNTQAGLST YVSAP AIIQ QIVALQSDLS SLQSDLENSL PDDRNRCINE LCTHIQNLQQ LLFASSTTAQ PILTPWPLMK ELDEMGKINS KLSTAV EEV TLEHRNKREI VKHHAKDVEL QRRVFVDFFC NPERLRNQVR ELNALVRARQ ASSS UniProtKB: AUGMIN subunit 3 |
-Macromolecule #3: AUGMIN subunit 5
| Macromolecule | Name: AUGMIN subunit 5 / type: protein_or_peptide / ID: 3 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: |
| Molecular weight | Theoretical: 84.325758 KDa |
| Recombinant expression | Organism: |
| Sequence | String: MQSLSSSAPT PEAILEWLQK EMGYRQLGPY NGSSKSHVPS IDAIRKICRG NMIPIWNFLI NRVKSEKTVE RIRRNITVHG GSSNASIGS SVNPGKEESK SKGRRKDKTV TGESSSYAED REAALQEREL AAKEVERLRN IVRRQRKDLK ARMLEVSREE A ERKRMLDE ...String: MQSLSSSAPT PEAILEWLQK EMGYRQLGPY NGSSKSHVPS IDAIRKICRG NMIPIWNFLI NRVKSEKTVE RIRRNITVHG GSSNASIGS SVNPGKEESK SKGRRKDKTV TGESSSYAED REAALQEREL AAKEVERLRN IVRRQRKDLK ARMLEVSREE A ERKRMLDE RANYRHKQAL LEAYDQQCDE ATRIFAEYHK RLQVYVNQAN DAQRSVNSSN EVLSSLSANS EREAVYSTVK GT KSADDVI LMETTRERNI RIVCDLLASR MIERIRNSFP AYEGNGICSL PELETAKLGF EYDGEITDEM KTVIVNSLRG PPL LLQAIA AYTLRIKTLI SREMEKIDVR ADAEMLRYKF ENNRVTDNSS SDVSSPSNNQ LLERQKAHVQ QFLATEDALN KAAE ARDLC HKFINRLHGS ADTATHSFVG GTTQSGSNLR QFELDVWGKE REAAGLRASL NTLLSEIQRL NKLCAERKEA EDSLK KKWK KIEEFDARRS ELETIYTTLL KANMDAVAFW NQQPLAAREY ASATVIPASE VVVDISNSAK DFIEKEVSAF FQSPDN SLY MLPATPQGLA RDPSAIPSIC RISAALQYPA GLEGSDASLA SVLESLEFCL RVRGSEACVL EDLAKAIDLV HIRQDLV ES GHSLLDHAFR AQQKYERTTN YCLDLASEQE NTISDQWLPE LRTAVQNAQA SSEHCKYVRG LLDEWWEQPA STVVDWVT V DGQSVAAWQN HVKQLLAFYD KESLRT UniProtKB: AUGMIN subunit 5 |
-Macromolecule #4: AUGMIN subunit 6
| Macromolecule | Name: AUGMIN subunit 6 / type: protein_or_peptide / ID: 4 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: |
| Molecular weight | Theoretical: 43.303605 KDa |
| Recombinant expression | Organism: |
| Sequence | String: MTMDREKERE LELESAMYTN CLLLGLDPNV IGLGASNGTP RVGLFRHSNP KLGEQLLYFI LSSLRGPAQS SKDFDKVWPI FDSAQSRDF RKVVQAIISE LESQGALPRS NSRVSSLATC CGPRFVELLW QLSLHALREV HRRTFPADVA SNPLPSSLTD V SFSHAATL ...String: MTMDREKERE LELESAMYTN CLLLGLDPNV IGLGASNGTP RVGLFRHSNP KLGEQLLYFI LSSLRGPAQS SKDFDKVWPI FDSAQSRDF RKVVQAIISE LESQGALPRS NSRVSSLATC CGPRFVELLW QLSLHALREV HRRTFPADVA SNPLPSSLTD V SFSHAATL LPVTKARIVL ERRRFLKNAE TAVQRQAMWS NLAHEMTAEF RGLCAEEAYL QQELEKLNDL RNKVKQEGEV WD DLVSSSS QNSHLVSKAT RLWDSIMARK GQHEVLASGP IEDLIAHREH RYRISGSALL AAMDQSSQVP RAELLSAHSD DSA SLADDK ELSDGSYTNM HDHSLVDSFE TASSQASDET LSRVDDRGGK INQTVDVAEV IRRWTHALQR I UniProtKB: AUGMIN subunit 6 |
-Macromolecule #5: AUGMIN subunit 7
| Macromolecule | Name: AUGMIN subunit 7 / type: protein_or_peptide / ID: 5 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: |
| Molecular weight | Theoretical: 37.046961 KDa |
| Recombinant expression | Organism: |
| Sequence | String: MAAKQMEEIQ KKLRLLSYPR ANAPAQSLLF AGMERYALLE WLFFKLLGDK SPFSQQNLQG DAGVRDEETV RIQYLAEIAK FLGITPTVD IEAIQGHGTY EDRMEMLRNI VDLVEASLFS DNQEWSIDEQ VAKDIQLIDA IAERQSLIFS EECKLFPADV Q IQSIYPLP ...String: MAAKQMEEIQ KKLRLLSYPR ANAPAQSLLF AGMERYALLE WLFFKLLGDK SPFSQQNLQG DAGVRDEETV RIQYLAEIAK FLGITPTVD IEAIQGHGTY EDRMEMLRNI VDLVEASLFS DNQEWSIDEQ VAKDIQLIDA IAERQSLIFS EECKLFPADV Q IQSIYPLP DVSELETKLS EQAKILSNLQ QKVDDLAAKH AYNPDEEYTE VESQLRARLE SFLETARAFN TIYTKEIRPW TH MMEVPQL HGFGPAANRL LEAYNMLLKF LGNLKNLRDS HAALSIGSSG TVAGEPSSVT RIVSDCEAAL TVLNRDLGIL SAS IAREQG ERL UniProtKB: AUGMIN subunit 7 |
-Macromolecule #6: AUGMIN subunit 8
| Macromolecule | Name: AUGMIN subunit 8 / type: protein_or_peptide / ID: 6 / Number of copies: 1 / Enantiomer: LEVO |
|---|---|
| Source (natural) | Organism: |
| Molecular weight | Theoretical: 31.372709 KDa |
| Recombinant expression | Organism: |
| Sequence | String: MKSSEDQVDP RLIDGKGSGR PSTPPSRGIS PSRIRQTTTS TQSSTTTSVL SFITDVKKGK KASYIEDVHQ LRLLHNRYLQ WRFAIARAE SVMYIQRLTS EETLFNVWHA ISELQDHVTR QRIGLQQLKL EIKLNSLLND QMVSLEDWAT LERDHVSSLV G AISDLEAN ...String: MKSSEDQVDP RLIDGKGSGR PSTPPSRGIS PSRIRQTTTS TQSSTTTSVL SFITDVKKGK KASYIEDVHQ LRLLHNRYLQ WRFAIARAE SVMYIQRLTS EETLFNVWHA ISELQDHVTR QRIGLQQLKL EIKLNSLLND QMVSLEDWAT LERDHVSSLV G AISDLEAN TLRLPATGGT KADTESLKAA MSSALDVMQA MGSSIWSLLS KVEEMNIMVT ELAVVVTKES SMQGKCEDLL AS TAIMQIE ECSLRTHLIQ TRREEGEDAE TPPPLLPLSK FPWP UniProtKB: AUGMIN subunit 8 |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.5 |
|---|---|
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy
Electron microscopy
| Microscope | TFS GLACIOS |
|---|---|
| Software | Name: SerialEM |
| Image recording | Film or detector model: GATAN K3 (6k x 4k) / Average electron dose: 60.0 e/Å2 |
| Electron beam | Acceleration voltage: 200 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD / Nominal defocus max: 1.8 µm / Nominal defocus min: 0.6 µm |

