 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | CryoEM structure of GNE-1952-alkylated KRAS G12C in complex with engineered conformationally rigid Fab 2H11.4DS | |||||||||
 マップデータ マップデータ | ||||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | fab / antigen binding fragment / protein engineering / gtpase / ONCOPROTEIN-MMUNE SYSTEM complex / ONCOPROTEIN-IMMUNE SYSTEM complex | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報response to mineralocorticoid / GMP binding / forebrain astrocyte development / LRR domain binding / regulation of synaptic transmission, GABAergic / negative regulation of epithelial cell differentiation / response to isolation stress / response to gravity / epithelial tube branching involved in lung morphogenesis / type I pneumocyte differentiation ...response to mineralocorticoid / GMP binding / forebrain astrocyte development / LRR domain binding / regulation of synaptic transmission, GABAergic / negative regulation of epithelial cell differentiation / response to isolation stress / response to gravity / epithelial tube branching involved in lung morphogenesis / type I pneumocyte differentiation / Rac protein signal transduction / positive regulation of Rac protein signal transduction / Signaling by RAS GAP mutants / Signaling by RAS GTPase mutants / Activation of RAS in B cells / myoblast proliferation / skeletal muscle cell differentiation / RAS signaling downstream of NF1 loss-of-function variants / RUNX3 regulates p14-ARF / positive regulation of glial cell proliferation / SOS-mediated signalling / Activated NTRK3 signals through RAS / Activated NTRK2 signals through RAS / SHC1 events in ERBB4 signaling / cardiac muscle cell proliferation / Signalling to RAS / Activated NTRK2 signals through FRS2 and FRS3 / SHC-related events triggered by IGF1R / Estrogen-stimulated signaling through PRKCZ / glial cell proliferation / SHC-mediated cascade:FGFR3 / MET activates RAS signaling / SHC-mediated cascade:FGFR2 / PTK6 Regulates RHO GTPases, RAS GTPase and MAP kinases / Signaling by PDGFRA transmembrane, juxtamembrane and kinase domain mutants / Signaling by PDGFRA extracellular domain mutants / SHC-mediated cascade:FGFR4 / Erythropoietin activates RAS / SHC-mediated cascade:FGFR1 / Signaling by FGFR4 in disease / Signaling by CSF3 (G-CSF) / FRS-mediated FGFR3 signaling / Signaling by FLT3 ITD and TKD mutants / FRS-mediated FGFR2 signaling / FRS-mediated FGFR4 signaling / p38MAPK events / FRS-mediated FGFR1 signaling / Signaling by FGFR3 in disease / protein-membrane adaptor activity / Tie2 Signaling / striated muscle cell differentiation / Signaling by FGFR2 in disease / GRB2 events in EGFR signaling / SHC1 events in EGFR signaling / Signaling by FLT3 fusion proteins / FLT3 Signaling / Signaling by FGFR1 in disease / EGFR Transactivation by Gastrin / NCAM signaling for neurite out-growth / homeostasis of number of cells within a tissue / CD209 (DC-SIGN) signaling / GRB2 events in ERBB2 signaling / Downstream signal transduction / Insulin receptor signalling cascade / Ras activation upon Ca2+ influx through NMDA receptor / SHC1 events in ERBB2 signaling / response to glucocorticoid / Constitutive Signaling by Overexpressed ERBB2 / Signaling by phosphorylated juxtamembrane, extracellular and kinase domain KIT mutants / VEGFR2 mediated cell proliferation / small monomeric GTPase / FCERI mediated MAPK activation / liver development / RAF activation / Signaling by ERBB2 TMD/JMD mutants / female pregnancy / Signaling by SCF-KIT / Signaling by high-kinase activity BRAF mutants / Constitutive Signaling by EGFRvIII / regulation of long-term neuronal synaptic plasticity / MAP2K and MAPK activation / Signaling by ERBB2 ECD mutants / Signaling by ERBB2 KD Mutants / visual learning / cytoplasmic side of plasma membrane / cytokine-mediated signaling pathway / Regulation of RAS by GAPs / Signaling by CSF1 (M-CSF) in myeloid cells / RAS processing / Negative regulation of MAPK pathway / Signaling by RAF1 mutants / Signaling by moderate kinase activity BRAF mutants / Paradoxical activation of RAF signaling by kinase inactive BRAF / Signaling downstream of RAS mutants / GDP binding / Signaling by BRAF and RAF1 fusions / positive regulation of cellular senescence / DAP12 signaling / MAPK cascade / Constitutive Signaling by Ligand-Responsive EGFR Cancer Variants 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.8 Å | |||||||||
 データ登録者 データ登録者 | Kung JE / Sudhamsu J | |||||||||
| 資金援助 | 1件
| |||||||||
 引用 引用 | ジャーナル: bioRxiv / 年: 2024 タイトル: Disulfi de constrained Fabs overcome target size limitation for high-resolution single-particle cryo-EM. 著者: Jennifer E Kung / Matthew C Johnson / Christine C Jao / Christopher P Arthur / Dimitry Tegunov / Alexis Rohou / Jawahar Sudhamsu /  要旨: High-resolution structures of proteins are critical to understanding molecular mechanisms of biological processes and in the discovery of therapeutic molecules. Cryo-EM has revolutionized structure ...High-resolution structures of proteins are critical to understanding molecular mechanisms of biological processes and in the discovery of therapeutic molecules. Cryo-EM has revolutionized structure determination of large proteins and their complexes, but a vast majority of proteins that underlie human diseases are small (< 50 kDa) and usually beyond its reach due to low signal-to-noise images and difficulties in particle alignment. Current strategies to overcome this problem increase the overall size of small protein targets using scaffold proteins that bind to the target, but are limited by inherent flexibility and not being bound to their targets in a rigid manner, resulting in the target being poorly resolved compared to the scaffolds. Here we present an iteratively engineered molecular design for transforming Fabs (antibody fragments), into conformationally rigid scaffolds (Rigid-Fabs) that, when bound to small proteins (~20 kDa), can enable high-resolution structure determination using cryo-EM. This design introduces multiple disulfide bonds at strategic locations, generates a well-folded Fab constrained into a rigid conformation and can be applied to Fabs from various species, isotypes and chimeric Fabs. We present examples of the Rigid Fab design enabling high-resolution (2.3-2.5 Å) structures of small proteins, Ang2 (26 kDa) and KRAS (21 kDa) by cryo-EM. The strategies for designing disulfide constrained Rigid Fabs in our work thus establish a general approach to overcome the target size limitation of single particle cryo-EM. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 添付画像 |
|---|

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_43221.map.gz | 21.6 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-43221-v30.xmlemd-43221.xml | 27.2 KB 27.2 KB | 表示 表示 | EMDBヘッダ |
| 画像 |  emd_43221.png emd_43221.png | 25.2 KB | ||
| マスクデータ | emd_43221_msk_1.map | 42.9 MB | マスクマップ | |
| Filedesc metadata | emd-43221.cif.gz | 6.9 KB | ||
| その他 | emd_43221_additional_1.map.gzemd_43221_additional_2.map.gzemd_43221_additional_3.map.gzemd_43221_half_map_1.map.gzemd_43221_half_map_2.map.gz | 39.8 MB 39.8 MB 21.7 MB 39.8 MB 39.8 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-43221ftp://ftp.pdbj.org/pub/emdb/structures/EMD-43221 http://ftp.pdbj.org/pub/emdb/structures/EMD-43221ftp://ftp.pdbj.org/pub/emdb/structures/EMD-43221 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_43221_validation.pdf.gz | 681.2 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_43221_full_validation.pdf.gz | 680.8 KB | 表示 | |
| XML形式データ | emd_43221_validation.xml.gz | 11.4 KB | 表示 | |
| CIF形式データ | emd_43221_validation.cif.gz | 13.5 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-43221ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-43221 | HTTPS FTP |
-関連構造データ
| 関連構造データ | 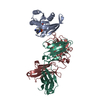 8vgqMC  8vegC  8vgeC  8vgfC  8vggC  8vghC 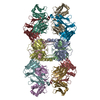 8vgiC 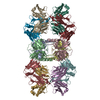 8vgjC 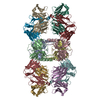 8vgkC  8vglC  8vgmC 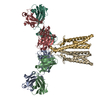 8vgnC 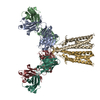 8vgoC  8vgpC M: このマップから作成された原子モデル C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |





















-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |
















-マップ
| ファイル | ダウンロード / ファイル: emd_43221.map.gz / 形式: CCP4 / 大きさ: 42.9 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.0573 Å | ||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
-マスク #1
| ファイル | emd_43221_msk_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-追加マップ: Half map for local refinement of KRAS G12C and Fab variable domain
| ファイル | emd_43221_additional_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Half map for local refinement of KRAS G12C and Fab variable domain | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-追加マップ: Half map for local refinement of KRAS G12C and Fab variable domain
| ファイル | emd_43221_additional_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Half map for local refinement of KRAS G12C and Fab variable domain | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-追加マップ: Local refinement map for KRAS G12C and Fab variable domain
| ファイル | emd_43221_additional_3.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | Local refinement map for KRAS G12C and Fab variable domain | ||||||||||||
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-ハーフマップ: #2
| ファイル | emd_43221_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








-ハーフマップ: #1
| ファイル | emd_43221_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
| 密度ヒストグラム |








- 試料の構成要素
試料の構成要素
+全体 : GNE-1952-alkylated KRAS G12C in complex with Fab 2H11.4DS
+超分子 #1: GNE-1952-alkylated KRAS G12C in complex with Fab 2H11.4DS
+超分子 #2: GNE-1952-alkylated KRAS G12C
+超分子 #3: Fab 2H11.4DS
+分子 #1: GTPase KRas

+分子 #2: Fab 2H11.4DS heavy chain

 Cricetulus griseus (モンゴルキヌゲネズミ)
Cricetulus griseus (モンゴルキヌゲネズミ)+分子 #3: Fab 2H11.4DS light chain
+分子 #4: 1-{4-[(7M)-6-methyl-7-(5-methyl-2H-indazol-4-yl)quinazolin-4-yl]p...
+分子 #5: GUANOSINE-5'-DIPHOSPHATE
+分子 #6: MAGNESIUM ION
+分子 #7: water

-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7 構成要素:
| |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| グリッド | モデル: Quantifoil R0.6/1 / 材質: GOLD / メッシュ: 300 / 支持フィルム - 材質: CARBON / 支持フィルム - トポロジー: HOLEY / 前処理 - タイプ: GLOW DISCHARGE 詳細: The peak fraction was subjected to mild crosslinking with 0.5 mM BS3 at room temperature for 10 min. The crosslinking reaction was quenched by addition of 100mM Tris pH 7.5. | |||||||||||||||
| 凍結 | 凍結剤: ETHANE |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: FEI FALCON IV (4k x 4k) 平均電子線量: 40.3 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | C2レンズ絞り径: 70.0 µm / 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.7 mm / 最大 デフォーカス(公称値): 1.5 µm / 最小 デフォーカス(公称値): 0.5 µm / 倍率(公称値): 105000 |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: NONE |
|---|---|
| 最終 再構成 | 想定した対称性 - 点群: C1 (非対称) / 解像度のタイプ: BY AUTHOR / 解像度: 2.8 Å / 解像度の算出法: FSC 0.143 CUT-OFF / ソフトウェア - 名称: cryoSPARC / 使用した粒子像数: 926738 |
| 初期 角度割当 | タイプ: MAXIMUM LIKELIHOOD / ソフトウェア - 名称: cryoSPARC |
| 最終 角度割当 | タイプ: OTHER / ソフトウェア - 名称: cryoSPARC |

