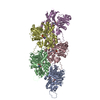
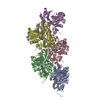
ジャーナル: Nat Struct Mol Biol / 年: 2023 タイトル: Molecular mechanisms of inorganic-phosphate release from the core and barbed end of actin filaments. 著者: Wout Oosterheert / Florian E C Blanc / Ankit Roy / Alexander Belyy / Micaela Boiero Sanders / Oliver Hofnagel / Gerhard Hummer / Peter Bieling / Stefan Raunser / 要旨: The release of inorganic phosphate (P) from actin filaments constitutes a key step in their regulated turnover, which is fundamental to many cellular functions. The mechanisms underlying P release ...The release of inorganic phosphate (P) from actin filaments constitutes a key step in their regulated turnover, which is fundamental to many cellular functions. The mechanisms underlying P release from the core and barbed end of actin filaments remain unclear. Here, using human and bovine actin isoforms, we combine cryo-EM with molecular-dynamics simulations and in vitro reconstitution to demonstrate how actin releases P through a 'molecular backdoor'. While constantly open at the barbed end, the backdoor is predominantly closed in filament-core subunits and opens only transiently through concerted amino acid rearrangements. This explains why P escapes rapidly from the filament end but slowly from internal subunits. In a nemaline-myopathy-associated actin variant, the backdoor is predominantly open in filament-core subunits, resulting in accelerated P release and filaments with drastically shortened ADP-P caps. Our results provide the molecular basis for P release from actin and exemplify how a disease-linked mutation distorts the nucleotide-state distribution and atomic structure of the filament.
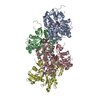
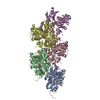
超分子 #1: Actin filament harboring the N111S mutation.
超分子
名称: Actin filament harboring the N111S mutation. / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1 詳細: Beta-actin was expressed as fusion protein, with thymosin beta4 and a deca-His-tag fused to the actin C-terminus. During the purification, thymosin beta-4 and the deca-His-tag were removed. ...詳細: Beta-actin was expressed as fusion protein, with thymosin beta4 and a deca-His-tag fused to the actin C-terminus. During the purification, thymosin beta-4 and the deca-His-tag were removed. Actin was purified as monomer from insect cells. It was then polymerized into a filament in vitro.
想定した対称性 - 点群: C1 (非対称) / 解像度のタイプ: BY AUTHOR / 解像度: 2.3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / ソフトウェア - 名称: RELION (ver. 3.1.0) 詳細: The final refinement was performed from local searches in RELION. 使用した粒子像数: 1756928
初期 角度割当
タイプ: MAXIMUM LIKELIHOOD / ソフトウェア - 名称: SPHIRE (ver. 1.4) 詳細: First 3D refinement was performed in helical SPHIRE with meridien alpha, which imposes helical restraints to limit particle shifts to the helical rise to prevent particle duplication, but ...詳細: First 3D refinement was performed in helical SPHIRE with meridien alpha, which imposes helical restraints to limit particle shifts to the helical rise to prevent particle duplication, but does not apply helical symmetry
最終 角度割当
タイプ: MAXIMUM LIKELIHOOD / ソフトウェア - 名称: RELION (ver. 3.1.0)
最終 3次元分類
クラス数: 8 / ソフトウェア - 名称: RELION (ver. 3.1.0) 詳細: 3D classification without image alignment was performed in RELION to remove particles that did not contain high-resolution information.
Chain - Source name: PDB / Chain - Initial model type: experimental model
詳細
chain C of pdb 8A2T (including all water molecules) was fit in the central actin subunit of the density map. After substitution of all alpha-actin specific amino-acids to the corresponding beta-actin residues, introducing the N111S mutation, and further manual model building in Coot, the resulting model was fitted in four more actin subunits (chains A, B, D, E) in the density map. The filament was modeled as a pentamer to capture the full interaction interface of the central subunit with its four neighboring subunits. All water molecules were first manually built, inspected and adjusted in the central subunit, and were then copied to the other chains with non-crystallographic symmetry (NCS). Because the local resolution was worse at the periphery of the reconstruction, we removed water molecules that displayed poor corresponding cryo-EM density in the non-central actin chains. The model was refined in Phenix real-space refine with NCS restraints but without Ramachandran and rotamer restraints.
精密化
空間: REAL / プロトコル: FLEXIBLE FIT
得られたモデル
PDB-8oid: Cryo-EM structure of ADP-bound, filamentous beta-actin harboring the N111S mutation
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報
 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 ドイツ, European Union, 3件
ドイツ, European Union, 3件  引用
引用 構造の表示
構造の表示
 ダウンロードとリンク
ダウンロードとリンク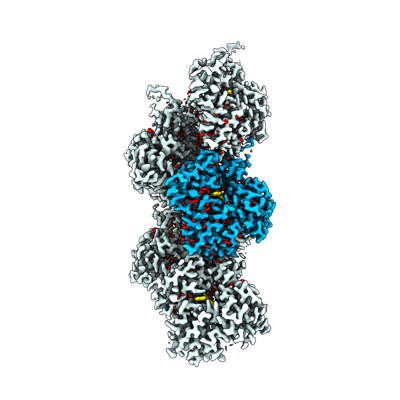 emd_16889.png
emd_16889.png http://ftp.pdbj.org/pub/emdb/structures/EMD-16889
http://ftp.pdbj.org/pub/emdb/structures/EMD-16889


















 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)













































 試料の構成要素
試料の構成要素 Trichoplusia ni (イラクサキンウワバ)
Trichoplusia ni (イラクサキンウワバ)

 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN