 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | DNA-PK in the active state | |||||||||
 Map data Map data | ||||||||||
 Sample Sample |
| |||||||||
 Keywords Keywords | DNA-dependent protein kinase / DNA BINDING PROTEIN | |||||||||
| Function / homology |  Function and homology information Function and homology informationpositive regulation of platelet formation / DNA-dependent protein kinase complex / Ku70:Ku80 complex / negative regulation of t-circle formation / DNA-dependent protein kinase activity / DNA end binding / small-subunit processome assembly / positive regulation of lymphocyte differentiation / histone H2AXS139 kinase activity / DNA-dependent protein kinase-DNA ligase 4 complex ...positive regulation of platelet formation / DNA-dependent protein kinase complex / Ku70:Ku80 complex / negative regulation of t-circle formation / DNA-dependent protein kinase activity / DNA end binding / small-subunit processome assembly / positive regulation of lymphocyte differentiation / histone H2AXS139 kinase activity / DNA-dependent protein kinase-DNA ligase 4 complex / immunoglobulin V(D)J recombination / nonhomologous end joining complex / cellular response to X-ray / regulation of epithelial cell proliferation / regulation of smooth muscle cell proliferation / double-strand break repair via classical nonhomologous end joining / double-strand break repair via alternative nonhomologous end joining / Cytosolic sensors of pathogen-associated DNA / nuclear telomere cap complex / telomere capping / IRF3-mediated induction of type I IFN / regulation of hematopoietic stem cell differentiation / U3 snoRNA binding / regulation of telomere maintenance / recombinational repair / cellular hyperosmotic salinity response / maturation of 5.8S rRNA / protein localization to chromosome, telomeric region / positive regulation of double-strand break repair via nonhomologous end joining / negative regulation of cGAS/STING signaling pathway / 2-LTR circle formation / peptidyl-threonine phosphorylation / telomeric repeat DNA binding / negative regulation of protein phosphorylation / DNA 3'-5' helicase / 5'-deoxyribose-5-phosphate lyase activity / 3'-5' DNA helicase activity / ATP-dependent activity, acting on DNA / telomere maintenance via telomerase / mitotic G1 DNA damage checkpoint signaling / positive regulation of erythrocyte differentiation / activation of innate immune response / telomere maintenance / cyclin binding / DNA helicase activity / protein modification process / DNA-(apurinic or apyrimidinic site) lyase / class I DNA-(apurinic or apyrimidinic site) endonuclease activity / site of DNA damage / positive regulation of translation / peptidyl-serine phosphorylation / intrinsic apoptotic signaling pathway in response to DNA damage / small-subunit processome / Nonhomologous End-Joining (NHEJ) / cellular response to gamma radiation / protein-DNA complex / regulation of circadian rhythm / double-strand break repair via nonhomologous end joining / enzyme activator activity / cellular response to insulin stimulus / rhythmic process / double-strand break repair / E3 ubiquitin ligases ubiquitinate target proteins / transcription regulator complex / scaffold protein binding / double-stranded DNA binding / DNA recombination / secretory granule lumen / ficolin-1-rich granule lumen / damaged DNA binding / RNA polymerase II-specific DNA-binding transcription factor binding / protein phosphorylation / protein kinase activity / chromosome, telomeric region / non-specific serine/threonine protein kinase / transcription cis-regulatory region binding / ribonucleoprotein complex / protein domain specific binding / innate immune response / protein serine kinase activity / negative regulation of DNA-templated transcription / protein serine/threonine kinase activity / ubiquitin protein ligase binding / Neutrophil degranulation / DNA damage response / nucleolus / negative regulation of apoptotic process / positive regulation of DNA-templated transcription / chromatin / protein-containing complex binding / enzyme binding / positive regulation of transcription by RNA polymerase II / ATP hydrolysis activity / protein-containing complex / DNA-templated transcription / DNA binding / RNA binding / extracellular region / nucleoplasm / ATP binding Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||
| Method | single particle reconstruction / cryo EM / Resolution: 2.91 Å | |||||||||
 Authors Authors | Liang S / Blundell TL | |||||||||
| Funding support |  United Kingdom, 1 items United Kingdom, 1 items
| |||||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2023 Title: Human DNA-dependent protein kinase activation mechanism. Authors: Shikang Liang / Tom L Blundell / Abstract: DNA-dependent protein kinase (DNA-PK), a multicomponent complex including the DNA-PK catalytic subunit and Ku70/80 heterodimer together with DNA, is central to human DNA damage response and repair. ...DNA-dependent protein kinase (DNA-PK), a multicomponent complex including the DNA-PK catalytic subunit and Ku70/80 heterodimer together with DNA, is central to human DNA damage response and repair. Using a DNA-PK-selective inhibitor (M3814), we identified from one dataset two cryo-EM structures of the human DNA-PK complex in different states, the intermediate state and the active state. Here we show that activation of the kinase is regulated through conformational changes caused by the binding ligand and the string region (residues 802-846) of the DNA-PK catalytic subunit, particularly the helix-hairpin-helix motif (residues 816-836) that interacts with DNA. These observations demonstrate the regulatory role of the ligand and explain why DNA-PK is DNA dependent. Cooperation and coordination among binding partners, disordered flexible regions and mechanically flexible HEAT repeats modulate the activation of the kinase. Together with previous findings, these results provide a better molecular understanding of DNA-PK catalysis. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Supplemental images |
|---|

- Downloads & links
Downloads & links
-EMDB archive
| Map data | emd_14545.map.gz | 6.9 MB | EMDB map data format | |
|---|---|---|---|---|
| Header (meta data) | emd-14545-v30.xmlemd-14545.xml | 22.3 KB 22.3 KB | Display Display | EMDB header |
| Images |  emd_14545.png emd_14545.png | 27.3 KB | ||
| Filedesc metadata | emd-14545.cif.gz | 9.3 KB | ||
| Others | emd_14545_half_map_1.map.gzemd_14545_half_map_2.map.gz | 151.8 MB 151.8 MB | ||
| Archive directory |  http://ftp.pdbj.org/pub/emdb/structures/EMD-14545ftp://ftp.pdbj.org/pub/emdb/structures/EMD-14545 http://ftp.pdbj.org/pub/emdb/structures/EMD-14545ftp://ftp.pdbj.org/pub/emdb/structures/EMD-14545 | HTTPS FTP |
-Related structure data
| Related structure data |  7z87MC  7z88C M: atomic model generated by this map C: citing same article ( |
|---|---|
| Similar structure data |

-Links
| EMDB pages | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| Related items in Molecule of the Month |









-Map
| File | Download / File: emd_14545.map.gz / Format: CCP4 / Size: 163.6 MB / Type: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & slices | Image control
Images are generated by Spider. | ||||||||||||||||||||||||||||||||||||
| Voxel size | X=Y=Z: 1.304 Å | ||||||||||||||||||||||||||||||||||||
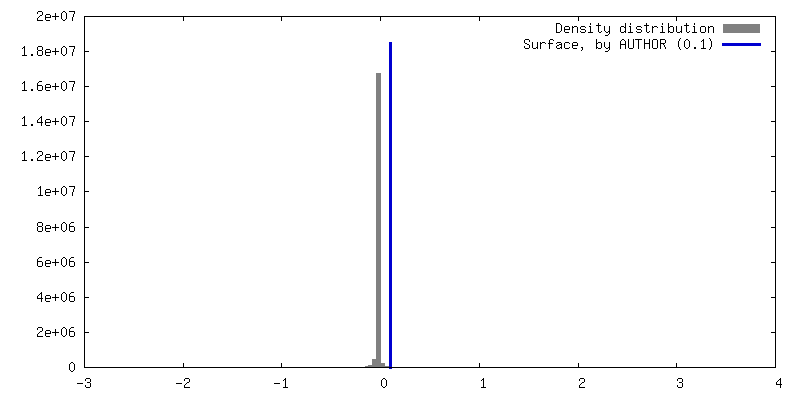
| Density |
| ||||||||||||||||||||||||||||||||||||
| Symmetry | Space group: 1 | ||||||||||||||||||||||||||||||||||||
| Details | EMDB XML:
|
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)



















-Supplemental data
-Half map: #2
| File | emd_14545_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
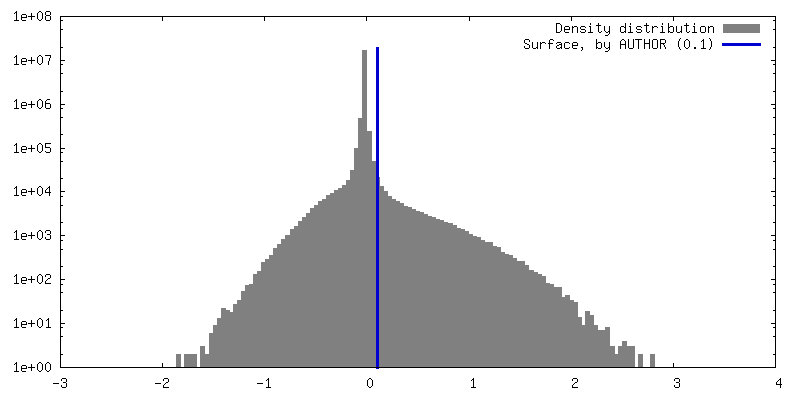
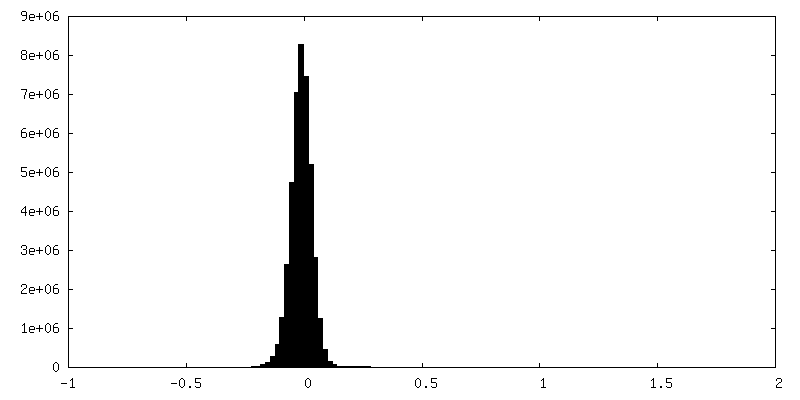
| Density Histograms |





-Half map: #1
| File | emd_14545_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Projections & Slices |
| ||||||||||||
| Density Histograms |



- Sample components
Sample components
-Entire : Human DNA-dependent protein kinase
| Entire | Name: Human DNA-dependent protein kinase |
|---|---|
| Components |
|
-Supramolecule #1: Human DNA-dependent protein kinase
| Supramolecule | Name: Human DNA-dependent protein kinase / type: complex / ID: 1 / Parent: 0 / Macromolecule list: #1-#5 |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
-Macromolecule #1: DNA-dependent protein kinase catalytic subunit
| Macromolecule | Name: DNA-dependent protein kinase catalytic subunit / type: protein_or_peptide / ID: 1 / Number of copies: 1 / Enantiomer: LEVO / EC number: non-specific serine/threonine protein kinase |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 469.673219 KDa |
| Sequence | String: MAGSGAGVRC SLLRLQETLS AADRCGAALA GHQLIRGLGQ ECVLSSSPAV LALQTSLVFS RDFGLLVFVR KSLNSIEFRE CREEILKFL CIFLEKMGQK IAPYSVEIKN TCTSVYTKDR AAKCKIPALD LLIKLLQTFR SSRLMDEFKI GELFSKFYGE L ALKKKIPD ...String: MAGSGAGVRC SLLRLQETLS AADRCGAALA GHQLIRGLGQ ECVLSSSPAV LALQTSLVFS RDFGLLVFVR KSLNSIEFRE CREEILKFL CIFLEKMGQK IAPYSVEIKN TCTSVYTKDR AAKCKIPALD LLIKLLQTFR SSRLMDEFKI GELFSKFYGE L ALKKKIPD TVLEKVYELL GLLGEVHPSE MINNAENLFR AFLGELKTQM TSAVREPKLP VLAGCLKGLS SLLCNFTKSM EE DPQTSRE IFNFVLKAIR PQIDLKRYAV PSAGLRLFAL HASQFSTCLL DNYVSLFEVL LKWCAHTNVE LKKAALSALE SFL KQVSNM VAKNAEMHKN KLQYFMEQFY GIIRNVDSNN KELSIAIRGY GLFAGPCKVI NAKDVDFMYV ELIQRCKQMF LTQT DTGDD RVYQMPSFLQ SVASVLLYLD TVPEVYTPVL EHLVVMQIDS FPQYSPKMQL VCCRAIVKVF LALAAKGPVL RNCIS TVVH QGLIRICSKP VVLPKGPESE SEDHRASGEV RTGKWKVPTY KDYVDLFRHL LSSDQMMDSI LADEAFFSVN SSSESL NHL LYDEFVKSVL KIVEKLDLTL EIQTVGEQEN GDEAPGVWMI PTSDPAANLH PAKPKDFSAF INLVEFCREI LPEKQAE FF EPWVYSFSYE LILQSTRLPL ISGFYKLLSI TVRNAKKIKY FEGVSPKSLK HSPEDPEKYS CFALFVKFGK EVAVKMKQ Y KDELLASCLT FLLSLPHNII ELDVRAYVPA LQMAFKLGLS YTPLAEVGLN ALEEWSIYID RHVMQPYYKD ILPCLDGYL KTSALSDETK NNWEVSALSR AAQKGFNKVV LKHLKKTKNL SSNEAISLEE IRIRVVQMLG SLGGQINKNL LTVTSSDEMM KSYVAWDRE KRLSFAVPFR EMKPVIFLDV FLPRVTELAL TASDRQTKVA ACELLHSMVM FMLGKATQMP EGGQGAPPMY Q LYKRTFPV LLRLACDVDQ VTRQLYEPLV MQLIHWFTNN KKFESQDTVA LLEAILDGIV DPVDSTLRDF CGRCIREFLK WS IKQITPQ QQEKSPVNTK SLFKRLYSLA LHPNAFKRLG ASLAFNNIYR EFREEESLVE QFVFEALVIY MESLALAHAD EKS LGTIQQ CCDAIDHLCR IIEKKHVSLN KAKKRRLPRG FPPSASLCLL DLVKWLLAHC GRPQTECRHK SIELFYKFVP LLPG NRSPN LWLKDVLKEE GVSFLINTFE GGGCGQPSGI LAQPTLLYLR GPFSLQATLC WLDLLLAALE CYNTFIGERT VGALQ VLGT EAQSSLLKAV AFFLESIAMH DIIAAEKCFG TGAAGNRTSP QEGERYNYSK CTVVVRIMEF TTTLLNTSPE GWKLLK KDL CNTHLMRVLV QTLCEPASIG FNIGDVQVMA HLPDVCVNLM KALKMSPYKD ILETHLREKI TAQSIEELCA VNLYGPD AQ VDRSRLAAVV SACKQLHRAG LLHNILPSQS TDLHHSVGTE LLSLVYKGIA PGDERQCLPS LDLSCKQLAS GLLELAFA F GGLCERLVSL LLNPAVLSTA SLGSSQGSVI HFSHGEYFYS LFSETINTEL LKNLDLAVLE LMQSSVDNTK MVSAVLNGM LDQSFRERAN QKHQGLKLAT TILQHWKKCD SWWAKDSPLE TKMAVLALLA KILQIDSSVS FNTSHGSFPE VFTTYISLLA DTKLDLHLK GQAVTLLPFF TSLTGGSLEE LRRVLEQLIV AHFPMQSREF PPGTPRFNNY VDCMKKFLDA LELSQSPMLL E LMTEVLCR EQQHVMEELF QSSFRRIARR GSCVTQVGLL ESVYEMFRKD DPRLSFTRQS FVDRSLLTLL WHCSLDALRE FF STIVVDA IDVLKSRFTK LNESTFDTQI TKKMGYYKIL DVMYSRLPKD DVHAKESKIN QVFHGSCITE GNELTKTLIK LCY DAFTEN MAGENQLLER RRLYHCAAYN CAISVICCVF NELKFYQGFL FSEKPEKNLL IFENLIDLKR RYNFPVEVEV PMER KKKYI EIRKEAREAA NGDSDGPSYM SSLSYLADST LSEEMSQFDF STGVQSYSYS SQDPRPATGR FRRREQRDPT VHDDV LELE MDELNRHECM APLTALVKHM HRSLGPPQGE EDSVPRDLPS WMKFLHGKLG NPIVPLNIRL FLAKLVINTE EVFRPY AKH WLSPLLQLAA SENNGGEGIH YMVVEIVATI LSWTGLATPT GVPKDEVLAN RLLNFLMKHV FHPKRAVFRH NLEIIKT LV ECWKDCLSIP YRLIFEKFSG KDPNSKDNSV GIQLLGIVMA NDLPPYDPQC GIQSSEYFQA LVNNMSFVRY KEVYAAAA E VLGLILRYVM ERKNILEESL CELVAKQLKQ HQNTMEDKFI VCLNKVTKSF PPLADRFMNA VFFLLPKFHG VLKTLCLEV VLCRVEGMTE LYFQLKSKDF VQVMRHRDDE RQKVCLDIIY KMMPKLKPVE LRELLNPVVE FVSHPSTTCR EQMYNILMWI HDNYRDPES ETDNDSQEIF KLAKDVLIQG LIDENPGLQL IIRNFWSHET RLPSNTLDRL LALNSLYSPK IEVHFLSLAT N FLLEMTSM SPDYPNPMFE HPLSECEFQE YTIDSDWRFR STVLTPMFVE TQASQGTLQT RTQEGSLSAR WPVAGQIRAT QQ QHDFTLT QTADGRSSFD WLTGSSTDPL VDHTSPSSDS LLFAHKRSER LQRAPLKSVG PDFGKKRLGL PGDEVDNKVK GAA GRTDLL RLRRRFMRDQ EKLSLMYARK GVAEQKREKE IKSELKMKQD AQVVLYRSYR HGDLPDIQIK HSSLITPLQA VAQR DPIIA KQLFSSLFSG ILKEMDKFKT LSEKNNITQK LLQDFNRFLN TTFSFFPPFV SCIQDISCQH AALLSLDPAA VSAGC LASL QQPVGIRLLE EALLRLLPAE LPAKRVRGKA RLPPDVLRWV ELAKLYRSIG EYDVLRGIFT SEIGTKQITQ SALLAE ARS DYSEAAKQYD EALNKQDWVD GEPTEAEKDF WELASLDCYN HLAEWKSLEY CSTASIDSEN PPDLNKIWSE PFYQETY LP YMIRSKLKLL LQGEADQSLL TFIDKAMHGE LQKAILELHY SQELSLLYLL QDDVDRAKYY IQNGIQSFMQ NYSSIDVL L HQSRLTKLQS VQALTEIQEF ISFISKQGNL SSQVPLKRLL NTWTNRYPDA KMDPMNIWDD IITNRCFFLS KIEEKLTPL PEDNSMNVDQ DGDPSDRMEV QEQEEDISSL IRSCKFSMKM KMIDSARKQN NFSLAMKLLK ELHKESKTRD DWLVSWVQSY CRLSHCRSR SQGCSEQVLT VLKTVSLLDE NNVSSYLSKN ILAFRDQNIL LGTTYRIIAN ALSSEPACLA EIEEDKARRI L ELSGSSSE DSEKVIAGLY QRAFQHLSEA VQAAEEEAQP PSWSCGPAAG VIDAYMTLAD FCDQQLRKEE ENASVIDSAE LQ AYPALVV EKMLKALKLN SNEARLKFPR LLQIIERYPE ETLSLMTKEI SSVPCWQFIS WISHMVALLD KDQAVAVQHS VEE ITDNYP QAIVYPFIIS SESYSFKDTS TGHKNKEFVA RIKSKLDQGG VIQDFINALD QLSNPELLFK DWSNDVRAEL AKTP VNKKN IEKMYERMYA ALGDPKAPGL GAFRRKFIQT FGKEFDKHFG KGGSKLLRMK LSDFNDITNM LLLKMNKDSK PPGNL KECS PWMSDFKVEF LRNELEIPGQ YDGRGKPLPE YHVRIAGFDE RVTVMASLRR PKRIIIRGHD EREHPFLVKG GEDLRQ DQR VEQLFQVMNG ILAQDSACSQ RALQLRTYSV VPMTSRLGLI EWLENTVTLK DLLLNTMSQE EKAAYLSDPR APPCEYK DW LTKMSGKHDV GAYMLMYKGA NRTETVTSFR KRESKVPADL LKRAFVRMST SPEAFLALRS HFASSHALIC ISHWILGI G DRHLNNFMVA METGGVIGID FGHAFGSATQ FLPVPELMPF RLTRQFINLM LPMKETGLMY SIMVHALRAF RSDPGLLTN TMDVFVKEPS FDWKNFEQKM LKKGGSWIQE INVAEKNWYP RQKICYAKRK LAGANPAVIT CDELLLGHEK APAFRDYVAV ARGSKDHNI RAQEPESGLS EETQVKCLMD QATDPNILGR TWEGWEPWM UniProtKB: DNA-dependent protein kinase catalytic subunit |
-Macromolecule #2: X-ray repair cross-complementing protein 6
| Macromolecule | Name: X-ray repair cross-complementing protein 6 / type: protein_or_peptide / ID: 2 / Number of copies: 1 / Enantiomer: LEVO EC number: Hydrolases; Acting on acid anhydrides; Acting on acid anhydrides to facilitate cellular and subcellular movement |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 69.945039 KDa |
| Recombinant expression | Organism:   Spodoptera frugiperda (fall armyworm) Spodoptera frugiperda (fall armyworm) |
| Sequence | String: MSGWESYYKT EGDEEAEEEQ EENLEASGDY KYSGRDSLIF LVDASKAMFE SQSEDELTPF DMSIQCIQSV YISKIISSDR DLLAVVFYG TEKDKNSVNF KNIYVLQELD NPGAKRILEL DQFKGQQGQK RFQDMMGHGS DYSLSEVLWV CANLFSDVQF K MSHKRIML ...String: MSGWESYYKT EGDEEAEEEQ EENLEASGDY KYSGRDSLIF LVDASKAMFE SQSEDELTPF DMSIQCIQSV YISKIISSDR DLLAVVFYG TEKDKNSVNF KNIYVLQELD NPGAKRILEL DQFKGQQGQK RFQDMMGHGS DYSLSEVLWV CANLFSDVQF K MSHKRIML FTNEDNPHGN DSAKASRART KAGDLRDTGI FLDLMHLKKP GGFDISLFYR DIISIAEDED LRVHFEESSK LE DLLRKVR AKETRKRALS RLKLKLNKDI VISVGIYNLV QKALKPPPIK LYRETNEPVK TKTRTFNTST GGLLLPSDTK RSQ IYGSRQ IILEKEETEE LKRFDDPGLM LMGFKPLVLL KKHHYLRPSL FVYPEESLVI GSSTLFSALL IKCLEKEVAA LCRY TPRRN IPPYFVALVP QEEELDDQKI QVTPPGFQLV FLPFADDKRK MPFTEKIMAT PEQVGKMKAI VEKLRFTYRS DSFEN PVLQ QHFRNLEALA LDLMEPEQAV DLTLPKVEAM NKRLGSLVDE FKELVYPPDY NPEGKVTKRK HDNEGSGSKR PKVEYS EEE LKTHISKGTL GKFTVPMLKE ACRAYGLKSG LKKQELLEAL TKHFQD UniProtKB: DNA repair protein Ku70 |
-Macromolecule #3: X-ray repair cross-complementing protein 5
| Macromolecule | Name: X-ray repair cross-complementing protein 5 / type: protein_or_peptide / ID: 3 / Number of copies: 1 / Enantiomer: LEVO EC number: Hydrolases; Acting on acid anhydrides; Acting on acid anhydrides to facilitate cellular and subcellular movement |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 82.812438 KDa |
| Recombinant expression | Organism: Spodoptera frugiperda (fall armyworm) |
| Sequence | String: MVRSGNKAAV VLCMDVGFTM SNSIPGIESP FEQAKKVITM FVQRQVFAEN KDEIALVLFG TDGTDNPLSG GDQYQNITVH RHLMLPDFD LLEDIESKIQ PGSQQADFLD ALIVSMDVIQ HETIGKKFEK RHIEIFTDLS SRFSKSQLDI IIHSLKKCDI S LQFFLPFS ...String: MVRSGNKAAV VLCMDVGFTM SNSIPGIESP FEQAKKVITM FVQRQVFAEN KDEIALVLFG TDGTDNPLSG GDQYQNITVH RHLMLPDFD LLEDIESKIQ PGSQQADFLD ALIVSMDVIQ HETIGKKFEK RHIEIFTDLS SRFSKSQLDI IIHSLKKCDI S LQFFLPFS LGKEDGSGDR GDGPFRLGGH GPSFPLKGIT EQQKEGLEIV KMVMISLEGE DGLDEIYSFS ESLRKLCVFK KI ERHSIHW PCRLTIGSNL SIRIAAYKSI LQERVKKTWT VVDAKTLKKE DIQKETVYCL NDDDETEVLK EDIIQGFRYG SDI VPFSKV DEEQMKYKSE GKCFSVLGFC KSSQVQRRFF MGNQVLKVFA ARDDEAAAVA LSSLIHALDD LDMVAIVRYA YDKR ANPQV GVAFPHIKHN YECLVYVQLP FMEDLRQYMF SSLKNSKKYA PTEAQLNAVD ALIDSMSLAK KDEKTDTLED LFPTT KIPN PRFQRLFQCL LHRALHPREP LPPIQQHIWN MLNPPAEVTT KSQIPLSKIK TLFPLIEAKK KDQVTAQEIF QDNHED GPT AKKLKTEQGG AHFSVSSLAE GSVTSVGSVN PAENFRVLVK QKKASFEEAS NQLINHIEQF LDTNETPYFM KSIDCIR AF REEAIKFSEE QRFNNFLKAL QEKVEIKQLN HFWEIVVQDG ITLITKEEAS GSSVTAEEAK KFLAPKDKPS GDTAAVFE E GGDVDDLLDM I UniProtKB: DNA repair protein Ku80 |
-Macromolecule #4: DNA (26-MER)
| Macromolecule | Name: DNA (26-MER) / type: dna / ID: 4 / Number of copies: 1 / Classification: DNA |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 7.900079 KDa |
| Sequence | String: (DC)(DC)(DC)(DG)(DC)(DT)(DG)(DC)(DC)(DG) (DA)(DT)(DT)(DC)(DC)(DG)(DC)(DT)(DG)(DG) (DA)(DA)(DC)(DA)(DT)(DT) |
-Macromolecule #5: DNA (26-MER)
| Macromolecule | Name: DNA (26-MER) / type: dna / ID: 5 / Number of copies: 1 / Classification: DNA |
|---|---|
| Source (natural) | Organism: Homo sapiens (human) |
| Molecular weight | Theoretical: 8.078203 KDa |
| Sequence | String: (DA)(DA)(DT)(DG)(DT)(DT)(DC)(DC)(DA)(DG) (DC)(DG)(DG)(DA)(DA)(DT)(DC)(DG)(DG)(DC) (DA)(DG)(DC)(DG)(DG)(DG) |
-Macromolecule #6: (~{S})-[2-chloranyl-4-fluoranyl-5-(7-morpholin-4-ylquinazolin-4-y...
| Macromolecule | Name: (~{S})-[2-chloranyl-4-fluoranyl-5-(7-morpholin-4-ylquinazolin-4-yl)phenyl]-(6-methoxypyridazin-3-yl)methanol type: ligand / ID: 6 / Number of copies: 1 / Formula: 1IX |
|---|---|
| Molecular weight | Theoretical: 481.907 Da |
| Chemical component information |  ChemComp-1IX: |
-Macromolecule #7: water
| Macromolecule | Name: water / type: ligand / ID: 7 / Number of copies: 194 / Formula: HOH |
|---|---|
| Molecular weight | Theoretical: 18.015 Da |
| Chemical component information |  ChemComp-HOH: |
-Experimental details
-Structure determination
| Method | cryo EM |
|---|---|
 Processing Processing | single particle reconstruction |
| Aggregation state | particle |
-Sample preparation
| Buffer | pH: 7.6 |
|---|---|
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy
Electron microscopy
| Microscope | FEI TITAN KRIOS |
|---|---|
| Image recording | Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) / Average electron dose: 47.22 e/Å2 |
| Electron beam | Acceleration voltage: 300 kV / Electron source:  FIELD EMISSION GUN FIELD EMISSION GUN |
| Electron optics | Illumination mode: FLOOD BEAM / Imaging mode: BRIGHT FIELD / Nominal defocus max: 2.3000000000000003 µm / Nominal defocus min: 1.1 µm |
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
-Image processing
| Startup model | Type of model: OTHER Details: PDB entry 6ZHA and 7K1N were used as the startup model. |
|---|---|
| Final reconstruction | Resolution.type: BY AUTHOR / Resolution: 2.91 Å / Resolution method: FSC 0.143 CUT-OFF / Number images used: 275300 |
| Initial angle assignment | Type: MAXIMUM LIKELIHOOD |
| Final angle assignment | Type: MAXIMUM LIKELIHOOD |
