 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8ox1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
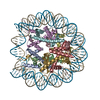
| タイトル | Structure of TRF1core in complex with telomeric nucleosome | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | DNA BINDING PROTEIN / Telomeric nucleosome / shelterin / telomere | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報positive regulation of shelterin complex assembly / negative regulation of establishment of protein localization to telomere / negative regulation of establishment of RNA localization to telomere / negative regulation of establishment of protein-containing complex localization to telomere / negative regulation of telomere maintenance via semi-conservative replication / negative regulation of telomeric D-loop disassembly / meiotic telomere clustering / t-circle formation / telomeric D-loop disassembly / shelterin complex ...positive regulation of shelterin complex assembly / negative regulation of establishment of protein localization to telomere / negative regulation of establishment of RNA localization to telomere / negative regulation of establishment of protein-containing complex localization to telomere / negative regulation of telomere maintenance via semi-conservative replication / negative regulation of telomeric D-loop disassembly / meiotic telomere clustering / t-circle formation / telomeric D-loop disassembly / shelterin complex / Telomere C-strand synthesis initiation / double-stranded telomeric DNA binding / ankyrin repeat binding / Telomere C-strand (Lagging Strand) Synthesis / nuclear telomere cap complex / G-rich strand telomeric DNA binding / telomere capping / Processive synthesis on the C-strand of the telomere / Polymerase switching on the C-strand of the telomere / Removal of the Flap Intermediate from the C-strand / DNA binding, bending / negative regulation of telomere maintenance via telomere lengthening / telomeric DNA binding / negative regulation of DNA replication / negative regulation of telomere maintenance via telomerase / positive regulation of telomere maintenance / Telomere Extension By Telomerase / telomere maintenance via telomerase / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / telomere organization / Interleukin-7 signaling / telomere maintenance / Inhibition of DNA recombination at telomere / RNA Polymerase I Promoter Opening / Meiotic synapsis / Assembly of the ORC complex at the origin of replication / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / SUMOylation of chromatin organization proteins / DNA methylation / Condensation of Prophase Chromosomes / epigenetic regulation of gene expression / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / innate immune response in mucosa / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / HDMs demethylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / HDACs deacetylate histones / RNA Polymerase I Promoter Escape / Nonhomologous End-Joining (NHEJ) / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / G2/M DNA damage checkpoint / Metalloprotease DUBs / NoRC negatively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / B-WICH complex positively regulates rRNA expression / PKMTs methylate histone lysines / Meiotic recombination / Pre-NOTCH Transcription and Translation / spindle / RMTs methylate histone arginines / Activation of anterior HOX genes in hindbrain development during early embryogenesis / fibrillar center / Transcriptional regulation of granulopoiesis / UCH proteinases / HCMV Early Events / antimicrobial humoral immune response mediated by antimicrobial peptide / structural constituent of chromatin / antibacterial humoral response / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / nucleosome / heterochromatin formation / RUNX1 regulates transcription of genes involved in differentiation of HSCs / nucleosome assembly / Processing of DNA double-strand break ends / HATs acetylate histones / Senescence-Associated Secretory Phenotype (SASP) / Factors involved in megakaryocyte development and platelet production / chromatin organization / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / Oxidative Stress Induced Senescence / microtubule binding 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.7 Å | ||||||||||||
 データ登録者 データ登録者 | Hu, H. / van Roon, A.M.M. / Ghanim, G.E. / Ahsan, B. / Oluwole, A. / Peak-Chew, S. / Robinson, C.V. / Nguyen, T.H.D. | ||||||||||||
| 資金援助 |  英国, European Union, 英国, European Union,  米国, 3件 米国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Sci Adv / 年: 2023 タイトル: Structural basis of telomeric nucleosome recognition by shelterin factor TRF1. 著者: Hongmiao Hu / Anne-Marie M van Roon / George E Ghanim / Bilal Ahsan / Abraham O Oluwole / Sew-Yeu Peak-Chew / Carol V Robinson / Thi Hoang Duong Nguyen / 要旨: Shelterin and nucleosomes are the key players that organize mammalian chromosome ends into the protective telomere caps. However, how they interact with each other at telomeres remains unknown. We ...Shelterin and nucleosomes are the key players that organize mammalian chromosome ends into the protective telomere caps. However, how they interact with each other at telomeres remains unknown. We report cryo-electron microscopy structures of a human telomeric nucleosome both unbound and bound to the shelterin factor TRF1. Our structures reveal that TRF1 binds unwrapped nucleosomal DNA ends by engaging both the nucleosomal DNA and the histone octamer. Unexpectedly, TRF1 binding shifts the register of the nucleosomal DNA by 1 bp. We discovered that phosphorylation of the TRF1 C terminus and a noncanomical DNA binding surface on TRF1 are critical for its association with telomeric nucleosomes. These insights into shelterin-chromatin interactions have crucial implications for understanding telomeric chromatin organization and other roles of shelterin at telomeres including replication and transcription. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8ox1.cif.gz | 367.5 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8ox1.ent.gz | 271 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8ox1.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8ox1_validation.pdf.gz | 1.4 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8ox1_full_validation.pdf.gz | 1.4 MB | 表示 | |
| XML形式データ | 8ox1_validation.xml.gz | 47.3 KB | 表示 | |
| CIF形式データ | 8ox1_validation.cif.gz | 74.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ox/8ox1ftp://data.pdbj.org/pub/pdb/validation_reports/ox/8ox1 | HTTPS FTP |
-関連構造データ


-リンク
 PDBj
PDBj












































- 集合体
集合体
| 登録構造単位 | 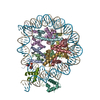
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 4種, 8分子 AEBFCGDH
| #1: タンパク質 | 分子量: 15805.581 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H3C1, H3FA, HIST1H3A, H3C2, H3FL, HIST1H3B, H3C3, H3FC HIST1H3C, H3C4, H3FB, HIST1H3D, H3C6, H3FD, HIST1H3E, H3C7, H3FI, HIST1H3F, H3C8, H3FH, HIST1H3G, H3C10, H3FK, HIST1H3H, H3C11, H3FF, ...遺伝子: H3C1, H3FA, HIST1H3A, H3C2, H3FL, HIST1H3B, H3C3, H3FC HIST1H3C, H3C4, H3FB, HIST1H3D, H3C6, H3FD, HIST1H3E, H3C7, H3FI, HIST1H3F, H3C8, H3FH, HIST1H3G, H3C10, H3FK, HIST1H3H, H3C11, H3FF, HIST1H3I, H3C12, H3FJ, HIST1H3J 発現宿主:  #2: タンパク質 | 分子量: 11762.839 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H4C1, H4/A, H4FA, HIST1H4A, H4C2, H4/I, H4FI, HIST1H4B, H4C3, H4/G, H4FG, HIST1H4C, H4C4, H4/B, H4FB, HIST1H4D, H4C5, H4/J, H4FJ, HIST1H4E, H4C6, H4/C, H4FC, HIST1H4F, H4C8, H4/H, H4FH, ...遺伝子: H4C1, H4/A, H4FA, HIST1H4A, H4C2, H4/I, H4FI, HIST1H4B, H4C3, H4/G, H4FG, HIST1H4C, H4C4, H4/B, H4FB, HIST1H4D, H4C5, H4/J, H4FJ, HIST1H4E, H4C6, H4/C, H4FC, HIST1H4F, H4C8, H4/H, H4FH, HIST1H4H, H4C9, H4/M, H4FM, HIST1H4I, H4C11, H4/E, H4FE, HIST1H4J, H4C12, H4/D, H4FD, HIST1H4K, H4C13, H4/K, H4FK, HIST1H4L, H4C14, H4/N, H4F2, H4FN, HIST2H4, HIST2H4A, H4C15, H4/O, H4FO, HIST2H4B, H4C16, H4-16, HIST4H4 発現宿主: #3: タンパク質 | 分子量: 14503.938 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: H2AC6, H2AFL, HIST1H2AC / 発現宿主: #4: タンパク質 | 分子量: 14305.627 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H2BC4, H2BFL, HIST1H2BC, H2BC6, H2BFH, HIST1H2BE, H2BC7, H2BFG, HIST1H2BF, H2BC8, H2BFA, HIST1H2BG, H2BC10, H2BFK, HIST1H2BI 発現宿主: |
|---|
-Telomeric DNA ... , 2種, 2分子 IJ
| #5: DNA鎖 | 分子量: 43478.914 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主: |
|---|---|
| #6: DNA鎖 | 分子量: 46024.258 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 発現宿主: |
-Telomeric repeat-binding factor ... , 2種, 2分子 LM
| #7: タンパク質 | 分子量: 50322.641 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: TERF1, PIN2, TRBF1, TRF, TRF1発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P54274 |
|---|---|
| #8: タンパク質 | 分子量: 50562.570 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: TERF1, PIN2, TRBF1, TRF, TRF1発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: P54274 |
-非ポリマー , 1種, 11分子 
| #9: 水 | ChemComp-HOH / 分子量: 18.015 Da / 分子数: 11 / 由来タイプ: 天然 / 式: H2O |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|---|
| Has protein modification | Y |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 |
| |||||||||||||||||||||||||||||||||||
| 由来(天然) |
| |||||||||||||||||||||||||||||||||||
| 由来(組換発現) |
| |||||||||||||||||||||||||||||||||||
| 緩衝液 | pH: 8 詳細: 25 mM HEPES-KOH pH 8.0, 150 mM KCl, 1 mM MgCl2, 1% glycerol, 0.01% Igepal CA-630, 1 mM DTT | |||||||||||||||||||||||||||||||||||
| 緩衝液成分 |
| |||||||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: C-flat-1.2/1.3 | |||||||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277.15 K / 詳細: Blot Force: -15 Blot Time: 2.5 s |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 105000 X / 最大 デフォーカス(公称値): 2500 nm / 最小 デフォーカス(公称値): 1000 nm / Cs: 2.7 mm / C2レンズ絞り径: 50 µm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER 最低温度: 78 K |
| 撮影 | 平均露光時間: 2.25 sec. / 電子線照射量: 56 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) / 撮影したグリッド数: 1 / 実像数: 24566 |
| 電子光学装置 | エネルギーフィルター名称: GIF Quantum LS / エネルギーフィルタースリット幅: 20 eV |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 画像処理 | 詳細: All images were processed using RELION4.0 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 5031654 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.7 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 93463 / クラス平均像の数: 1 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 2.7→125.55 Å / Cor.coef. Fo:Fc: 0.879 / SU B: 10.283 / SU ML: 0.19 / ESU R: 0.313 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | イオンプローブ半径: 0.8 Å / 減衰半径: 0.8 Å / VDWプローブ半径: 1.2 Å / 溶媒モデル: MASK | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 107.728 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 合計: 13191 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|