 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7uxh | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | cryo-EM structure of the mTORC1-TFEB-Rag-Ragulator complex | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | SIGNALING PROTEIN / mTORC1 / TFEB / Lysosome biogenesis / Autophagy | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報regulation of cholesterol import / positive regulation of protein localization to lysosome / regulation of cell-substrate junction organization / Gtr1-Gtr2 GTPase complex / regulation of cholesterol efflux / FNIP-folliculin RagC/D GAP / Ragulator complex / protein localization to cell junction / RNA polymerase III type 2 promoter sequence-specific DNA binding / RNA polymerase III type 1 promoter sequence-specific DNA binding ...regulation of cholesterol import / positive regulation of protein localization to lysosome / regulation of cell-substrate junction organization / Gtr1-Gtr2 GTPase complex / regulation of cholesterol efflux / FNIP-folliculin RagC/D GAP / Ragulator complex / protein localization to cell junction / RNA polymerase III type 2 promoter sequence-specific DNA binding / RNA polymerase III type 1 promoter sequence-specific DNA binding / positive regulation of cytoplasmic translational initiation / regulation of locomotor rhythm / T-helper 1 cell lineage commitment / positive regulation of pentose-phosphate shunt / positive regulation of wound healing, spreading of epidermal cells / TORC2 signaling / antibacterial innate immune response / regulation of TORC1 signaling / TORC2 complex / regulation of membrane permeability / cellular response to leucine starvation / negative regulation of lysosome organization / heart valve morphogenesis / TFIIIC-class transcription factor complex binding / protein localization to lysosome / TORC1 complex / voluntary musculoskeletal movement / positive regulation of transcription of nucleolar large rRNA by RNA polymerase I / positive regulation of odontoblast differentiation / calcineurin-NFAT signaling cascade / RNA polymerase III type 3 promoter sequence-specific DNA binding / positive regulation of keratinocyte migration / regulation of osteoclast differentiation / regulation of TOR signaling / regulation of lysosome organization / fibroblast migration / MTOR signalling / lysosome localization / cellular response to nutrient / cellular response to L-leucine / energy reserve metabolic process / Amino acids regulate mTORC1 / regulation of autophagosome assembly / Energy dependent regulation of mTOR by LKB1-AMPK / endosome organization / TORC1 signaling / serine/threonine protein kinase complex / ruffle organization / cellular response to methionine / positive regulation of osteoclast differentiation / negative regulation of cell size / positive regulation of ubiquitin-dependent protein catabolic process / kinase activator activity / cellular response to osmotic stress / negative regulation of protein localization to nucleus / anoikis / inositol hexakisphosphate binding / protein localization to membrane / cardiac muscle cell development / negative regulation of calcineurin-NFAT signaling cascade / regulation of myelination / endosomal transport / azurophil granule membrane / positive regulation of transcription by RNA polymerase III / lysosome organization / small GTPase-mediated signal transduction / negative regulation of macroautophagy / Macroautophagy / positive regulation of myotube differentiation / regulation of cell size / RHOJ GTPase cycle / Constitutive Signaling by AKT1 E17K in Cancer / positive regulation of actin filament polymerization / RHOQ GTPase cycle / germ cell development / TOR signaling / behavioral response to pain / mTORC1-mediated signalling / humoral immune response / oligodendrocyte differentiation / CDC42 GTPase cycle / positive regulation of oligodendrocyte differentiation / positive regulation of translational initiation / RHOH GTPase cycle / tertiary granule membrane / RHOG GTPase cycle / ficolin-1-rich granule membrane / protein serine/threonine kinase inhibitor activity / regulation of receptor recycling / social behavior / protein kinase activator activity / CD28 dependent PI3K/Akt signaling / RAC2 GTPase cycle / HSF1-dependent transactivation / RAC3 GTPase cycle / positive regulation of TOR signaling / regulation of macroautophagy / enzyme-substrate adaptor activity / positive regulation of G1/S transition of mitotic cell cycle / embryonic placenta development 類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.2 Å | ||||||
 データ登録者 データ登録者 | Cui, Z. / Hurley, J. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Nature / 年: 2023 タイトル: Structure of the lysosomal mTORC1-TFEB-Rag-Ragulator megacomplex. 著者: Zhicheng Cui / Gennaro Napolitano / Mariana E G de Araujo / Alessandra Esposito / Jlenia Monfregola / Lukas A Huber / Andrea Ballabio / James H Hurley /   要旨: The transcription factor TFEB is a master regulator of lysosomal biogenesis and autophagy. The phosphorylation of TFEB by the mechanistic target of rapamycin complex 1 (mTORC1) is unique in its ...The transcription factor TFEB is a master regulator of lysosomal biogenesis and autophagy. The phosphorylation of TFEB by the mechanistic target of rapamycin complex 1 (mTORC1) is unique in its mTORC1 substrate recruitment mechanism, which is strictly dependent on the amino acid-mediated activation of the RagC GTPase activating protein FLCN. TFEB lacks the TOR signalling motif responsible for the recruitment of other mTORC1 substrates. We used cryogenic-electron microscopy to determine the structure of TFEB as presented to mTORC1 for phosphorylation, which we refer to as the 'megacomplex'. Two full Rag-Ragulator complexes present each molecule of TFEB to the mTOR active site. One Rag-Ragulator complex is bound to Raptor in the canonical mode seen previously in the absence of TFEB. A second Rag-Ragulator complex (non-canonical) docks onto the first through a RagC GDP-dependent contact with the second Ragulator complex. The non-canonical Rag dimer binds the first helix of TFEB with a RagC-dependent aspartate clamp in the cleft between the Rag G domains. In cellulo mutation of the clamp drives TFEB constitutively into the nucleus while having no effect on mTORC1 localization. The remainder of the 108-amino acid TFEB docking domain winds around Raptor and then back to RagA. The double use of RagC GDP contacts in both Rag dimers explains the strong dependence of TFEB phosphorylation on FLCN and the RagC GDP state. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7uxh.cif.gz | 3.9 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7uxh.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 7uxh.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7uxh_validation.pdf.gz | 1.7 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7uxh_full_validation.pdf.gz | 1.7 MB | 表示 | |
| XML形式データ | 7uxh_validation.xml.gz | 259.9 KB | 表示 | |
| CIF形式データ | 7uxh_validation.cif.gz | 413.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/ux/7uxhftp://data.pdbj.org/pub/pdb/validation_reports/ux/7uxh | HTTPS FTP |
-関連構造データ










-リンク
 PDBj
PDBj






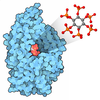












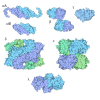


- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 4種, 8分子 ACBDEUTj
| #1: タンパク質 | 分子量: 289257.969 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MTOR, FRAP, FRAP1, FRAP2, RAFT1, RAPT1 / 発現宿主: Homo sapiens (ヒト)参照: UniProt: P42345, non-specific serine/threonine protein kinase #2: タンパク質 | 分子量: 35910.090 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: MLST8, GBL, LST8 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q9BVC4#3: タンパク質 | 分子量: 149200.016 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RPTOR, KIAA1303, RAPTOR / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q8N122#11: タンパク質 | 分子量: 52926.621 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: TFEB, BHLHE35 / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: P19484 |
|---|
-Ras-related GTP-binding protein ... , 2種, 8分子 FMVcGNWd
| #4: タンパク質 | 分子量: 36600.195 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RRAGA / 発現宿主: Homo sapiens (ヒト)参照: UniProt: Q7L523, 加水分解酵素; 酸無水物に作用; GTPに作用・細胞または細胞小器官の運動に関与 #5: タンパク質 | 分子量: 44298.859 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: RRAGC / 発現宿主: Homo sapiens (ヒト) / 参照: UniProt: Q9HB90 |
|---|
-Ragulator complex protein ... , 5種, 20分子 HOXeIPYfJQZgKRahLSbi
| #6: タンパク質 | 分子量: 17762.775 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR1, C11orf59, PDRO, PP7157発現宿主:   Spodoptera frugiperda (ツマジロクサヨトウ) Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q6IAA8 #7: タンパク質 | 分子量: 13517.450 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR2, MAPBPIP, ROBLD3, HSPC003発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9Y2Q5 #8: タンパク質 | 分子量: 13637.678 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR3, MAP2K1IP1, MAPKSP1, PRO2783発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q9UHA4 #9: タンパク質 | 分子量: 10753.236 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR4, C7orf59発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: Q0VGL1 #10: タンパク質 | 分子量: 9622.900 Da / 分子数: 4 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: LAMTOR5, HBXIP, XIP発現宿主: Spodoptera frugiperda (ツマジロクサヨトウ)参照: UniProt: O43504 |
|---|
-非ポリマー , 4種, 14分子 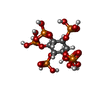
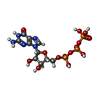

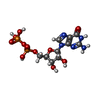
| #12: 化合物 |  分子量: 660.035 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C6H18O24P6 分子量: 660.035 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C6H18O24P6#13: 化合物 | ChemComp-GTP /  分子量: 523.180 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H16N5O14P3 / コメント: GTP, エネルギー貯蔵分子*YM 分子量: 523.180 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H16N5O14P3 / コメント: GTP, エネルギー貯蔵分子*YM#14: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 4 / 由来タイプ: 合成 / 式: Mg#15: 化合物 | ChemComp-GDP /  タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM タイプ: RNA linking / 分子量: 443.201 Da / 分子数: 4 / 由来タイプ: 合成 / 式: C10H15N5O11P2 / コメント: GDP, エネルギー貯蔵分子*YM |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: The mTORC1-TFEB-Rag-Ragulator complex / タイプ: COMPLEX / Entity ID: #1-#11 / 由来: MULTIPLE SOURCES |
|---|---|
| 緩衝液 | pH: 7.4 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2200 nm / 最小 デフォーカス(公称値): 800 nm |
| 撮影 | 電子線照射量: 50 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) |
- 解析
解析
| ソフトウェア |
| ||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.2 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 192332 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 101.66 Å2 | ||||||||||||||||||||||||||||||||
| 拘束条件 |
|
