 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 7u50 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | APE1 bound to a nucleosome core particle with AP-site at SHL-6 | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | DNA BINDING PROTEIN/DNA / nucleosome / DNA damage / DNA repair / DNA BINDING PROTEIN / DNA BINDING PROTEIN-DNA complex | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Resolution of Abasic Sites (AP sites) / phosphodiesterase activity, acting on 3'-phosphoglycolate-terminated DNA strands / telomere maintenance via base-excision repair / class II DNA-(apurinic or apyrimidinic site) endonuclease activity / : / DNA-(abasic site) binding / double-stranded DNA exodeoxyribonuclease activity / double-stranded telomeric DNA binding / double-stranded DNA 3'-5' DNA exonuclease activity / exodeoxyribonuclease III ...Resolution of Abasic Sites (AP sites) / phosphodiesterase activity, acting on 3'-phosphoglycolate-terminated DNA strands / telomere maintenance via base-excision repair / class II DNA-(apurinic or apyrimidinic site) endonuclease activity / : / DNA-(abasic site) binding / double-stranded DNA exodeoxyribonuclease activity / double-stranded telomeric DNA binding / double-stranded DNA 3'-5' DNA exonuclease activity / exodeoxyribonuclease III / Displacement of DNA glycosylase by APEX1 / 3'-5'-DNA exonuclease activity / positive regulation of gene expression via chromosomal CpG island demethylation / phosphoric diester hydrolase activity / 加水分解酵素; エステル加水分解酵素; 5'-リン酸モノエステル産生エンドデオキシリボヌクレアーゼ / uracil DNA N-glycosylase activity / DNA catabolic process / phosphodiesterase I activity / Resolution of AP sites via the multiple-nucleotide patch replacement pathway / Abasic sugar-phosphate removal via the single-nucleotide replacement pathway / POLB-Dependent Long Patch Base Excision Repair / PCNA-Dependent Long Patch Base Excision Repair / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Chromatin modifying enzymes / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / base-excision repair, gap-filling / regulation of mRNA stability / 3'-5' exonuclease activity / DNA-(apurinic or apyrimidinic site) endonuclease activity / Deposition of new CENPA-containing nucleosomes at the centromere / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / telomere organization / Interleukin-7 signaling / Inhibition of DNA recombination at telomere / RNA Polymerase I Promoter Opening / Meiotic synapsis / telomere maintenance / Assembly of the ORC complex at the origin of replication / cell redox homeostasis / SUMOylation of chromatin organization proteins / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / HCMV Late Events / SIRT1 negatively regulates rRNA expression / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / innate immune response in mucosa / DNA endonuclease activity / Regulation of endogenous retroelements by KRAB-ZFP proteins / Defective pyroptosis / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / HDACs deacetylate histones / Nonhomologous End-Joining (NHEJ) / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / Formation of the beta-catenin:TCF transactivating complex / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / HDMs demethylate histones / G2/M DNA damage checkpoint / NoRC negatively regulates rRNA expression / base-excision repair / DNA Damage/Telomere Stress Induced Senescence / B-WICH complex positively regulates rRNA expression / PKMTs methylate histone lysines / Meiotic recombination / chromatin DNA binding / Pre-NOTCH Transcription and Translation / Metalloprotease DUBs / RMTs methylate histone arginines / Activation of anterior HOX genes in hindbrain development during early embryogenesis / Transcriptional regulation of granulopoiesis / HCMV Early Events / RNA-DNA hybrid ribonuclease activity / antimicrobial humoral immune response mediated by antimicrobial peptide / structural constituent of chromatin / transcription corepressor activity / UCH proteinases / antibacterial humoral response / nucleosome / heterochromatin formation / nucleosome assembly / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / HATs acetylate histones / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Factors involved in megakaryocyte development and platelet production / chromatin organization / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / Processing of DNA double-strand break ends / Senescence-Associated Secretory Phenotype (SASP) / endonuclease activity 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト)synthetic construct (人工物) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.4 Å | |||||||||
 データ登録者 データ登録者 | Weaver, T.M. / Freudenthal, B.D. | |||||||||
| 資金援助 |  米国, 2件 米国, 2件
| |||||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2022 タイトル: Structural basis for APE1 processing DNA damage in the nucleosome. 著者: Tyler M Weaver / Nicole M Hoitsma / Jonah J Spencer / Lokesh Gakhar / Nicholas J Schnicker / Bret D Freudenthal / 要旨: Genomic DNA is continually exposed to endogenous and exogenous factors that promote DNA damage. Eukaryotic genomic DNA is packaged into nucleosomes, which present a barrier to accessing and ...Genomic DNA is continually exposed to endogenous and exogenous factors that promote DNA damage. Eukaryotic genomic DNA is packaged into nucleosomes, which present a barrier to accessing and effectively repairing DNA damage. The mechanisms by which DNA repair proteins overcome this barrier to repair DNA damage in the nucleosome and protect genomic stability is unknown. Here, we determine how the base excision repair (BER) endonuclease AP-endonuclease 1 (APE1) recognizes and cleaves DNA damage in the nucleosome. Kinetic assays determine that APE1 cleaves solvent-exposed AP sites in the nucleosome with 3 - 6 orders of magnitude higher efficiency than occluded AP sites. A cryo-electron microscopy structure of APE1 bound to a nucleosome containing a solvent-exposed AP site reveal that APE1 uses a DNA sculpting mechanism for AP site recognition, where APE1 bends the nucleosomal DNA to access the AP site. Notably, additional biochemical and structural characterization of occluded AP sites identify contacts between the nucleosomal DNA and histone octamer that prevent efficient processing of the AP site by APE1. These findings provide a rationale for the position-dependent activity of BER proteins in the nucleosome and suggests the ability of BER proteins to sculpt nucleosomal DNA drives efficient BER in chromatin. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7u50.cif.gz | 368.8 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7u50.ent.gz | 279.1 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 7u50.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7u50_validation.pdf.gz | 918.2 KB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7u50_full_validation.pdf.gz | 934.4 KB | 表示 | |
| XML形式データ | 7u50_validation.xml.gz | 40.4 KB | 表示 | |
| CIF形式データ | 7u50_validation.cif.gz | 64 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/u5/7u50ftp://data.pdbj.org/pub/pdb/validation_reports/u5/7u50 | HTTPS FTP |
-関連構造データ




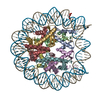
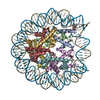
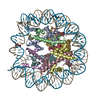
-リンク
 PDBj
PDBj












































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 5種, 9分子 AEBFCGDHK
| #1: タンパク質 | 分子量: 15257.838 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H3C15, HIST2H3A, H3C14, H3F2, H3FM, HIST2H3C, H3C13, HIST2H3D 発現宿主:  #2: タンパク質 | 分子量: 11263.231 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H4C1, H4/A, H4FA, HIST1H4A, H4C2, H4/I, H4FI, HIST1H4B, H4C3, H4/G, H4FG, HIST1H4C, H4C4, H4/B, H4FB, HIST1H4D, H4C5, H4/J, H4FJ, HIST1H4E, H4C6, H4/C, H4FC, HIST1H4F, H4C8, H4/H, H4FH, ...遺伝子: H4C1, H4/A, H4FA, HIST1H4A, H4C2, H4/I, H4FI, HIST1H4B, H4C3, H4/G, H4FG, HIST1H4C, H4C4, H4/B, H4FB, HIST1H4D, H4C5, H4/J, H4FJ, HIST1H4E, H4C6, H4/C, H4FC, HIST1H4F, H4C8, H4/H, H4FH, HIST1H4H, H4C9, H4/M, H4FM, HIST1H4I, H4C11, H4/E, H4FE, HIST1H4J, H4C12, H4/D, H4FD, HIST1H4K, H4C13, H4/K, H4FK, HIST1H4L, H4C14, H4/N, H4F2, H4FN, HIST2H4, HIST2H4A, H4C15, H4/O, H4FO, HIST2H4B, H4-16, HIST4H4 発現宿主: #3: タンパク質 | 分子量: 13990.342 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H2AC11, H2AFP, HIST1H2AG, H2AC13, H2AFC, HIST1H2AI, H2AC15, H2AFD, HIST1H2AK, H2AC16, H2AFI, HIST1H2AL, H2AC17, H2AFN, HIST1H2AM 発現宿主: #4: タンパク質 | 分子量: 13806.018 Da / 分子数: 2 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト)遺伝子: H2BC4, H2BFL, HIST1H2BC, H2BC6, H2BFH, HIST1H2BE, H2BC7, H2BFG, HIST1H2BF, H2BC8, H2BFA, HIST1H2BG, H2BC10, H2BFK, HIST1H2BI 発現宿主: #7: タンパク質 | | 分子量: 35606.492 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: APEX1, APE, APE1, APEX, APX, HAP1, REF1 / 発現宿主: 参照: UniProt: P27695, 加水分解酵素; エステル加水分解酵素 |
|---|
-DNA鎖 , 2種, 2分子 IJ
| #5: DNA鎖 | 分子量: 45005.656 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|---|
| #6: DNA鎖 | 分子量: 45610.043 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: APE1 bound to a nucleosome core particle with AP-site at SHL-6 タイプ: COMPLEX / Entity ID: all / 由来: RECOMBINANT | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||
| 由来(天然) | 生物種: Homo sapiens (ヒト) | ||||||||||||||||||||
| 由来(組換発現) | 生物種: | ||||||||||||||||||||
| 緩衝液 | pH: 7.4 | ||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | ||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 2200 nm / 最小 デフォーカス(公称値): 800 nm |
| 撮影 | 電子線照射量: 50 e/Å2 / フィルム・検出器のモデル: GATAN K3 (6k x 4k) |
- 解析
解析
| ソフトウェア | 名称: PHENIX / バージョン: 1.19.2_4158: / 分類: 精密化 | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.4 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 58854 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 拘束条件 |
|
