 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
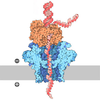
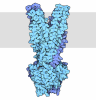
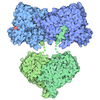
| 登録情報 | データベース: PDB / ID: 7pmn | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | S. cerevisiae replisome-SCF(Dia2) complex bound to double-stranded DNA (conformation II) | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | REPLICATION / Genome stability / DNA replication / Ubiquitination / termination / replisome / cryo-EM / CMG / SCF(Dia2) | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報RAVE complex / Iron uptake and transport / CBF3 complex / regulation of transcription by galactose / regulation of sulfur amino acid metabolic process / cellular response to methylmercury / establishment of sister chromatid cohesion / vacuolar proton-transporting V-type ATPase complex assembly / maintenance of DNA repeat elements / DNA-templated DNA replication maintenance of fidelity ...RAVE complex / Iron uptake and transport / CBF3 complex / regulation of transcription by galactose / regulation of sulfur amino acid metabolic process / cellular response to methylmercury / establishment of sister chromatid cohesion / vacuolar proton-transporting V-type ATPase complex assembly / maintenance of DNA repeat elements / DNA-templated DNA replication maintenance of fidelity / gene conversion / septin ring assembly / Unwinding of DNA / invasive growth in response to glucose limitation / replication fork arrest / Cul8-RING ubiquitin ligase complex / DNA replication initiation / protein-containing complex disassembly / meiotic chromosome segregation / epsilon DNA polymerase complex / MCM core complex / Assembly of the pre-replicative complex / Switching of origins to a post-replicative state / MCM complex binding / GINS complex / DNA strand elongation involved in mitotic DNA replication / nuclear DNA replication / mitotic DNA replication preinitiation complex assembly / premeiotic DNA replication / pre-replicative complex assembly involved in nuclear cell cycle DNA replication / mitotic DNA replication / regulation of exit from mitosis / SUMO binding / Activation of the pre-replicative complex / CMG complex / establishment of mitotic sister chromatid cohesion / DNA replication checkpoint signaling / kinetochore assembly / single-stranded 3'-5' DNA helicase activity / entrainment of circadian clock / single-stranded DNA 3'-5' DNA exonuclease activity / nuclear pre-replicative complex / regulation of metabolic process / MCM complex / Activation of ATR in response to replication stress / DNA replication preinitiation complex / Termination of translesion DNA synthesis / replication fork protection complex / mitotic DNA replication checkpoint signaling / mitotic DNA replication initiation / positive regulation of glucose transmembrane transport / protein neddylation / double-strand break repair via break-induced replication / mitotic intra-S DNA damage checkpoint signaling / vacuolar acidification / silent mating-type cassette heterochromatin formation / regulation of DNA-templated DNA replication initiation / 加水分解酵素; エステル加水分解酵素; 5'-リン酸モノエステル産生エンドデオキシリボヌクレアーゼ / exit from mitosis / mitochondrial fusion / single-stranded DNA helicase activity / SCF ubiquitin ligase complex / nucleotide-excision repair, DNA gap filling / DNA replication proofreading / mitotic sister chromatid cohesion / DNA strand elongation involved in DNA replication / SCF-dependent proteasomal ubiquitin-dependent protein catabolic process / nuclear chromosome / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / leading strand elongation / replication fork processing / Orc1 removal from chromatin / regulation of DNA replication / DNA unwinding involved in DNA replication / nuclear replication fork / 3'-5' DNA helicase activity / DNA replication origin binding / cullin family protein binding / Antigen processing: Ubiquitination & Proteasome degradation / Dual incision in TC-NER / subtelomeric heterochromatin formation / DNA replication initiation / regulation of protein-containing complex assembly / endomembrane system / negative regulation of cytoplasmic translation / error-prone translesion synthesis / heterochromatin formation / regulation of mitotic cell cycle / helicase activity / base-excision repair, gap-filling / meiotic cell cycle / replication fork / base-excision repair / DNA-templated DNA replication / kinetochore / double-strand break repair via nonhomologous end joining / G1/S transition of mitotic cell cycle / G2/M transition of mitotic cell cycle / double-strand break repair / nucleosome assembly 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  Synthetic construct (人工物) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.2 Å | ||||||||||||
 データ登録者 データ登録者 | Jenkyn-Bedford, M. / Yeeles, J.T.P. / Deegan, T.D. | ||||||||||||
| 資金援助 |  英国, 3件 英国, 3件
| ||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2021 タイトル: A conserved mechanism for regulating replisome disassembly in eukaryotes. 著者: Michael Jenkyn-Bedford / Morgan L Jones / Yasemin Baris / Karim P M Labib / Giuseppe Cannone / Joseph T P Yeeles / Tom D Deegan / 要旨: Replisome disassembly is the final step of eukaryotic DNA replication and is triggered by ubiquitylation of the CDC45-MCM-GINS (CMG) replicative helicase. Despite being driven by evolutionarily ...Replisome disassembly is the final step of eukaryotic DNA replication and is triggered by ubiquitylation of the CDC45-MCM-GINS (CMG) replicative helicase. Despite being driven by evolutionarily diverse E3 ubiquitin ligases in different eukaryotes (SCF in budding yeast, CUL2 in metazoa), replisome disassembly is governed by a common regulatory principle, in which ubiquitylation of CMG is suppressed before replication termination, to prevent replication fork collapse. Recent evidence suggests that this suppression is mediated by replication fork DNA. However, it is unknown how SCF and CUL2 discriminate terminated from elongating replisomes, to selectively ubiquitylate CMG only after termination. Here we used cryo-electron microscopy to solve high-resolution structures of budding yeast and human replisome-E3 ligase assemblies. Our structures show that the leucine-rich repeat domains of Dia2 and LRR1 are structurally distinct, but bind to a common site on CMG, including the MCM3 and MCM5 zinc-finger domains. The LRR-MCM interaction is essential for replisome disassembly and, crucially, is occluded by the excluded DNA strand at replication forks, establishing the structural basis for the suppression of CMG ubiquitylation before termination. Our results elucidate a conserved mechanism for the regulation of replisome disassembly in eukaryotes, and reveal a previously unanticipated role for DNA in preserving replisome integrity. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 7pmn.cif.gz | 1.7 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb7pmn.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 7pmn.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 7pmn_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 7pmn_full_validation.pdf.gz | 1.2 MB | 表示 | |
| XML形式データ | 7pmn_validation.xml.gz | 199.7 KB | 表示 | |
| CIF形式データ | 7pmn_validation.cif.gz | 324.2 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/pm/7pmnftp://data.pdbj.org/pub/pdb/validation_reports/pm/7pmn | HTTPS FTP |
-関連構造データ

























-リンク
 PDBj
PDBj

























































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA replication licensing factor ... , 5種, 5分子 23467
| #1: タンパク質 | 分子量: 98911.539 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM2, YBL023C, YBL0438 / 発現宿主: |
|---|---|
| #2: タンパク質 | 分子量: 111987.562 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM3, YEL032W, SYGP-ORF23 / 発現宿主: |
| #3: タンパク質 | 分子量: 105138.375 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM4, CDC54, HCD21, YPR019W, YP9531.13 / 発現宿主: |
| #5: タンパク質 | 分子量: 113110.211 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM6, YGL201C / 発現宿主: |
| #6: タンパク質 | 分子量: 95049.875 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM7, CDC47, YBR202W, YBR1441 / 発現宿主: |
-タンパク質 , 7種, 9分子 5EFGHKLXY
| #4: タンパク質 | 分子量: 86505.734 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MCM5, CDC46, YLR274W, L9328.1 / 発現宿主: | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| #11: タンパク質 | 分子量: 75154.703 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: CDC45, SLD4, YLR103C, L8004.11 / 発現宿主: | ||||||||
| #12: タンパク質 | 分子量: 108610.148 Da / 分子数: 3 / Fragment: Mcm6 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: CTF4, CHL15, POB1, YPR135W, P9659.7 / 発現宿主: #15: タンパク質 | | 分子量: 22357.270 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: SKP1, CBF3D, YDR328C, D9798.14 / 発現宿主: #16: タンパク質 | | 分子量: 85368.844 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: DIA2, YOR080W, YOR29-31 / 発現宿主: #19: タンパク質 | | 分子量: 141296.875 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: TOF1, YNL273W, N0636 / 発現宿主: #20: タンパク質 | | 分子量: 36588.758 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: CSM3, YMR048W, YM9796.01 / 発現宿主: |
-DNA replication complex GINS protein ... , 4種, 4分子 ABCD
| #7: タンパク質 | 分子量: 24230.576 Da / 分子数: 1 / Fragment: Tof1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PSF1, YDR013W, PZA208, YD8119.18 / 発現宿主: |
|---|---|
| #8: タンパク質 | 分子量: 25096.807 Da / 分子数: 1 / Fragment: Csm3 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PSF2, YJL072C, HRF213, J1086 / 発現宿主: |
| #9: タンパク質 | 分子量: 22004.160 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: PACBIOSEQ_LOCUS5150, PACBIOSEQ_LOCUS5239, PACBIOSEQ_LOCUS5244, PACBIOSEQ_LOCUS5270, PACBIOSEQ_LOCUS5331 発現宿主: |
| #10: タンパク質 | 分子量: 33983.617 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: SLD5, YDR489W / 発現宿主: |
-DNA鎖 , 2種, 2分子 IJ
| #13: DNA鎖 | 分子量: 37738.676 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) Synthetic construct (人工物) |
|---|---|
| #14: DNA鎖 | 分子量: 35259.238 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
-DNA polymerase epsilon ... , 2種, 2分子 QR
| #17: タンパク質 | 分子量: 255890.422 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: POL2, DUN2, YNL262W, N0825 / 発現宿主: 参照: UniProt: P21951, DNA-directed DNA polymerase, 加水分解酵素; エステル加水分解酵素; 5'-リン酸モノエステル産生エンドデオキシリボヌクレアーゼ |
|---|---|
| #18: タンパク質 | 分子量: 78425.852 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: DPB2, YPR175W, P9705.7 / 発現宿主: |
-非ポリマー , 3種, 11分子 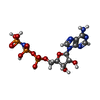


| #21: 化合物 |  分子量: 506.196 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H17N6O12P3 / タイプ: SUBJECT OF INVESTIGATION 分子量: 506.196 Da / 分子数: 2 / 由来タイプ: 合成 / 式: C10H17N6O12P3 / タイプ: SUBJECT OF INVESTIGATIONコメント: AMP-PNP, エネルギー貯蔵分子類似体*YM #22: 化合物 |  分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION 分子量: 24.305 Da / 分子数: 2 / 由来タイプ: 合成 / 式: Mg / タイプ: SUBJECT OF INVESTIGATION#23: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 7 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION 分子量: 65.409 Da / 分子数: 7 / 由来タイプ: 合成 / 式: Zn / タイプ: SUBJECT OF INVESTIGATION |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | Y |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | ||||||||||||||||||||||||||||
| 由来(天然) |
| ||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.6 | ||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||
| 試料支持 | 詳細: 15 mA / グリッドの材料: COPPER / グリッドのサイズ: 400 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | ||||||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE / 詳細: Manual plunger |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 81000 X / 最大 デフォーカス(公称値): 2200 nm / 最小 デフォーカス(公称値): 400 nm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 4 sec. / 電子線照射量: 38.8 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 実像数: 12730 |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum / エネルギーフィルタースリット幅: 20 eV |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 2160000 | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.2 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 369254 詳細: Composite map produced from individual cryo-EM density maps (resolutions 3.2 - 4.0 A) using Phenix combine_focused_maps. See publication for details. 対称性のタイプ: POINT |
