 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 6tu3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
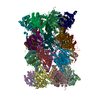
| タイトル | Rat 20S proteasome | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | HYDROLASE / PROTEASOME | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Cross-presentation of soluble exogenous antigens (endosomes) / SCF-beta-TrCP mediated degradation of Emi1 / Regulation of ornithine decarboxylase (ODC) / Assembly of the pre-replicative complex / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / Cdc20:Phospho-APC/C mediated degradation of Cyclin A ...Cross-presentation of soluble exogenous antigens (endosomes) / SCF-beta-TrCP mediated degradation of Emi1 / Regulation of ornithine decarboxylase (ODC) / Assembly of the pre-replicative complex / GSK3B and BTRC:CUL1-mediated-degradation of NFE2L2 / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / SCF(Skp2)-mediated degradation of p27/p21 / Autodegradation of the E3 ubiquitin ligase COP1 / Asymmetric localization of PCP proteins / Degradation of DVL / Hedgehog ligand biogenesis / Dectin-1 mediated noncanonical NF-kB signaling / Degradation of GLI1 by the proteasome / Hedgehog 'on' state / TNFR2 non-canonical NF-kB pathway / NIK-->noncanonical NF-kB signaling / UCH proteinases / CDK-mediated phosphorylation and removal of Cdc6 / G2/M Checkpoints / Ubiquitin Mediated Degradation of Phosphorylated Cdc25A / Ubiquitin-dependent degradation of Cyclin D / The role of GTSE1 in G2/M progression after G2 checkpoint / FBXL7 down-regulates AURKA during mitotic entry and in early mitosis / RUNX1 regulates transcription of genes involved in differentiation of HSCs / Regulation of RUNX3 expression and activity / GLI3 is processed to GLI3R by the proteasome / Activation of NF-kappaB in B cells / Degradation of beta-catenin by the destruction complex / Degradation of AXIN / Regulation of RAS by GAPs / Orc1 removal from chromatin / Neddylation / AUF1 (hnRNP D0) binds and destabilizes mRNA / MAPK6/MAPK4 signaling / Regulation of PTEN stability and activity / KEAP1-NFE2L2 pathway / Separation of Sister Chromatids / Antigen processing: Ubiquitination & Proteasome degradation / ABC-family proteins mediated transport / Ub-specific processing proteases / proteasome core complex / Neutrophil degranulation / immune system process / myofibril / NF-kappaB binding / proteasome endopeptidase complex / proteasome core complex, beta-subunit complex / proteasome core complex, alpha-subunit complex / threonine-type endopeptidase activity / negative regulation of inflammatory response to antigenic stimulus / : / sarcomere / proteasome complex / ciliary basal body / proteolysis involved in protein catabolic process / lipopolysaccharide binding / P-body / response to virus / response to organic cyclic compound / nuclear matrix / peptidase activity / positive regulation of NF-kappaB transcription factor activity / postsynapse / proteasome-mediated ubiquitin-dependent protein catabolic process / endopeptidase activity / response to oxidative stress / ribosome / centrosome / ubiquitin protein ligase binding / synapse / proteolysis / RNA binding / nucleoplasm / identical protein binding / nucleus / cytoplasm / cytosol 類似検索 - 分子機能 | |||||||||
| 生物種 |  | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.7 Å | |||||||||
 データ登録者 データ登録者 | Deshmukh, F.K. / Polkinghorn, C.R. / Elad, N. / Sharon, M. | |||||||||
| 資金援助 |  イスラエル, 2件 イスラエル, 2件
| |||||||||
 引用 引用 | ジャーナル: ACS Cent Sci / 年: 2020 タイトル: Comparative Structural Analysis of 20S Proteasome Ortholog Protein Complexes by Native Mass Spectrometry. 著者: Shay Vimer / Gili Ben-Nissan / David Morgenstern / Fanindra Kumar-Deshmukh / Caley Polkinghorn / Royston S Quintyn / Yury V Vasil'ev / Joseph S Beckman / Nadav Elad / Vicki H Wysocki / Michal Sharon /  要旨: Ortholog protein complexes are responsible for equivalent functions in different organisms. However, during evolution, each organism adapts to meet its physiological needs and the environmental ...Ortholog protein complexes are responsible for equivalent functions in different organisms. However, during evolution, each organism adapts to meet its physiological needs and the environmental challenges imposed by its niche. This selection pressure leads to structural diversity in protein complexes, which are often difficult to specify, especially in the absence of high-resolution structures. Here, we describe a multilevel experimental approach based on native mass spectrometry (MS) tools for elucidating the structural preservation and variations among highly related protein complexes. The 20S proteasome, an essential protein degradation machinery, served as our model system, wherein we examined five complexes isolated from different organisms. We show that throughout evolution, from the archaeal prokaryotic complex to the eukaryotic 20S proteasomes in yeast () and mammals (rat - , rabbit - and human - HEK293 cells), the proteasome increased both in size and stability. Native MS structural signatures of the rat and rabbit 20S proteasomes, which heretofore lacked high-resolution, three-dimensional structures, highly resembled that of the human complex. Using cryoelectron microscopy single-particle analysis, we were able to obtain a high-resolution structure of the rat 20S proteasome, allowing us to validate the MS-based results. Our study also revealed that the yeast complex, and not those in mammals, was the largest in size and displayed the greatest degree of kinetic stability. Moreover, we also identified a new proteoform of the PSMA7 subunit that resides within the rat and rabbit complexes, which to our knowledge have not been previously described. Altogether, our strategy enables elucidation of the unique structural properties of protein complexes that are highly similar to one another, a framework that is valid not only to ortholog protein complexes, but also for other highly related protein assemblies. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 6tu3.cif.gz | 1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb6tu3.ent.gz | 847.7 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 6tu3.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 6tu3_validation.pdf.gz | 1.5 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 6tu3_full_validation.pdf.gz | 1.6 MB | 表示 | |
| XML形式データ | 6tu3_validation.xml.gz | 167.2 KB | 表示 | |
| CIF形式データ | 6tu3_validation.cif.gz | 253.7 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/tu/6tu3ftp://data.pdbj.org/pub/pdb/validation_reports/tu/6tu3 | HTTPS FTP |
-関連構造データ
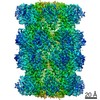

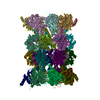

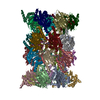


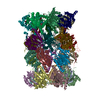


-リンク
 PDBj
PDBj









- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-Proteasome subunit alpha type- ... , 7種, 14分子 AOBPCQDRESFTGU
| #1: タンパク質 | 分子量: 27432.459 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #2: タンパク質 | 分子量: 25955.549 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #3: タンパク質 | 分子量: 29539.830 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #4: タンパク質 | 分子量: 28369.439 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #5: タンパク質 | 分子量: 26416.850 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #6: タンパク質 | 分子量: 29557.541 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #7: タンパク質 | 分子量: 28456.275 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-Proteasome subunit beta type- ... , 7種, 14分子 HVIWJXKYLZMaNb
| #8: タンパク質 | 分子量: 25309.576 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #9: タンパク質 | 分子量: 29963.461 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #10: タンパク質 | 分子量: 22988.895 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #11: タンパク質 | 分子量: 22941.381 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #12: タンパク質 | 分子量: 28615.408 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #13: タンパク質 | 分子量: 26511.301 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #14: タンパク質 | 分子量: 29226.248 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: Rat liver 20S proteasome / タイプ: COMPLEX 詳細: Endogenous 20S proteasome purified from the rat livers by anion-exchange chromatography Entity ID: all / 由来: NATURAL |
|---|---|
| 由来(天然) | 生物種: |
| 緩衝液 | pH: 7.4 |
| 緩衝液成分 | 濃度: 160 mM / 名称: Potassium phosphate / 式: K3PO4 |
| 試料 | 濃度: 14 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES / 詳細: Purified endogenous rat liver 20S proteasome |
| 試料支持 | 詳細: Grids were glow discharged 30 minutes before applying the sample. グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: C-flat-2/2 |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K / 詳細: blot for 3 seconds |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS 詳細: Preliminary grid screening was performed on Talos Arctica |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: OTHER |
| 電子レンズ | モード: DIFFRACTION / 倍率(公称値): 105000 X / 最大 デフォーカス(公称値): 1600 nm / 最小 デフォーカス(公称値): 600 nm / Cs: 2.7 mm / C2レンズ絞り径: 70 µm |
| 試料ホルダ | 凍結剤: NITROGEN 試料ホルダーモデル: FEI TITAN KRIOS AUTOGRID HOLDER |
| 撮影 | 平均露光時間: 1.5 sec. / 電子線照射量: 37 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) 撮影したグリッド数: 1 / 実像数: 2234 詳細: Images were collected in movie-mode at 45 frames per second |
| 電子光学装置 | エネルギーフィルター名称: GIF Bioquantum / エネルギーフィルタースリット幅: 20 eV |
- 解析
解析
| EMソフトウェア |
| ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 284161 | ||||||||||||||||||||||||||||
| 対称性 | 点対称性: C2 (2回回転対称) | ||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.7 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 245800 / アルゴリズム: BACK PROJECTION / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: FLEXIBLE FIT |
