Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 6rny | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
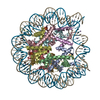
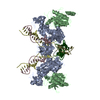
| Title | PFV intasome - nucleosome strand transfer complex | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | DNA BINDING PROTEIN / chromatin / nucleosome / retrovirus | |||||||||
| Function / homology |  Function and homology information Function and homology informationBarr body / negative regulation of chromosome condensation / pericentric heterochromatin formation / inner kinetochore / muscle cell differentiation / oocyte maturation / Hydrolases; Acting on peptide bonds (peptidases); Aspartic endopeptidases / ribonuclease H / nucleosomal DNA binding / nucleus organization ...Barr body / negative regulation of chromosome condensation / pericentric heterochromatin formation / inner kinetochore / muscle cell differentiation / oocyte maturation / Hydrolases; Acting on peptide bonds (peptidases); Aspartic endopeptidases / ribonuclease H / nucleosomal DNA binding / nucleus organization / spermatid development / single fertilization / subtelomeric heterochromatin formation / RNA polymerase II core promoter sequence-specific DNA binding / negative regulation of megakaryocyte differentiation / protein localization to CENP-A containing chromatin / Replacement of protamines by nucleosomes in the male pronucleus / CENP-A containing nucleosome / Packaging Of Telomere Ends / embryo implantation / Recognition and association of DNA glycosylase with site containing an affected purine / Cleavage of the damaged purine / Deposition of new CENPA-containing nucleosomes at the centromere / telomere organization / Recognition and association of DNA glycosylase with site containing an affected pyrimidine / Cleavage of the damaged pyrimidine / RNA Polymerase I Promoter Opening / Inhibition of DNA recombination at telomere / Assembly of the ORC complex at the origin of replication / Meiotic synapsis / Regulation of endogenous retroelements by the Human Silencing Hub (HUSH) complex / SUMOylation of chromatin organization proteins / DNA methylation / Condensation of Prophase Chromosomes / Chromatin modifications during the maternal to zygotic transition (MZT) / SIRT1 negatively regulates rRNA expression / HCMV Late Events / ERCC6 (CSB) and EHMT2 (G9a) positively regulate rRNA expression / PRC2 methylates histones and DNA / Regulation of endogenous retroelements by KRAB-ZFP proteins / innate immune response in mucosa / Defective pyroptosis / HDACs deacetylate histones / Regulation of endogenous retroelements by Piwi-interacting RNAs (piRNAs) / Nonhomologous End-Joining (NHEJ) / RNA Polymerase I Promoter Escape / Transcriptional regulation by small RNAs / DNA integration / Formation of the beta-catenin:TCF transactivating complex / Activated PKN1 stimulates transcription of AR (androgen receptor) regulated genes KLK2 and KLK3 / HDMs demethylate histones / RUNX1 regulates genes involved in megakaryocyte differentiation and platelet function / viral genome integration into host DNA / G2/M DNA damage checkpoint / Negative Regulation of CDH1 Gene Transcription / establishment of integrated proviral latency / NoRC negatively regulates rRNA expression / RNA-directed DNA polymerase / PKMTs methylate histone lysines / B-WICH complex positively regulates rRNA expression / DNA Damage/Telomere Stress Induced Senescence / virion component / male gonad development / Meiotic recombination / Pre-NOTCH Transcription and Translation / viral penetration into host nucleus / RNA-directed DNA polymerase activity / Activation of anterior HOX genes in hindbrain development during early embryogenesis / multicellular organism growth / RNA-DNA hybrid ribonuclease activity / Transcriptional regulation of granulopoiesis / RMTs methylate histone arginines / Transferases; Transferring phosphorus-containing groups; Nucleotidyltransferases / Metalloprotease DUBs / HCMV Early Events / osteoblast differentiation / structural constituent of chromatin / UCH proteinases / nucleosome / antimicrobial humoral immune response mediated by antimicrobial peptide / host cell / heterochromatin formation / nucleosome assembly / HATs acetylate histones / antibacterial humoral response / E3 ubiquitin ligases ubiquitinate target proteins / Recruitment and ATM-mediated phosphorylation of repair and signaling proteins at DNA double strand breaks / Factors involved in megakaryocyte development and platelet production / MLL4 and MLL3 complexes regulate expression of PPARG target genes in adipogenesis and hepatic steatosis / chromatin organization / RUNX1 regulates transcription of genes involved in differentiation of HSCs / positive regulation of cell growth / Processing of DNA double-strand break ends / Senescence-Associated Secretory Phenotype (SASP) / DNA recombination / Oxidative Stress Induced Senescence / Estrogen-dependent gene expression / DNA-directed DNA polymerase / aspartic-type endopeptidase activity / Hydrolases; Acting on ester bonds Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) Human spumaretrovirus Human spumaretrovirus Pyrobaculum filamentous virus 1 Pyrobaculum filamentous virus 1 | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.9 Å | |||||||||
 Authors Authors | Pye, V.E. / Renault, L. / Maskell, D.P. / Cherepanov, P. / Costa, A. | |||||||||
| Funding support |  United Kingdom, 2items United Kingdom, 2items
| |||||||||
 Citation Citation | Journal: Nat Commun / Year: 2019 Title: Retroviral integration into nucleosomes through DNA looping and sliding along the histone octamer. Authors: Marcus D Wilson / Ludovic Renault / Daniel P Maskell / Mohamed Ghoneim / Valerie E Pye / Andrea Nans / David S Rueda / Peter Cherepanov / Alessandro Costa /  Abstract: Retroviral integrase can efficiently utilise nucleosomes for insertion of the reverse-transcribed viral DNA. In face of the structural constraints imposed by the nucleosomal structure, integrase ...Retroviral integrase can efficiently utilise nucleosomes for insertion of the reverse-transcribed viral DNA. In face of the structural constraints imposed by the nucleosomal structure, integrase gains access to the scissile phosphodiester bonds by lifting DNA off the histone octamer at the site of integration. To clarify the mechanism of DNA looping by integrase, we determined a 3.9 Å resolution structure of the prototype foamy virus intasome engaged with a nucleosome core particle. The structural data along with complementary single-molecule Förster resonance energy transfer measurements reveal twisting and sliding of the nucleosomal DNA arm proximal to the integration site. Sliding the nucleosomal DNA by approximately two base pairs along the histone octamer accommodates the necessary DNA lifting from the histone H2A-H2B subunits to allow engagement with the intasome. Thus, retroviral integration into nucleosomes involves the looping-and-sliding mechanism for nucleosomal DNA repositioning, bearing unexpected similarities to chromatin remodelers. #1: Journal: Nat Commun / Year: 2019Title: Retroviral integration into nucleosomes through DNA looping and sliding along the histone octamer. Authors: Marcus D Wilson / Ludovic Renault / Daniel P Maskell / Mohamed Ghoneim / Valerie E Pye / Andrea Nans / David S Rueda / Peter Cherepanov / Alessandro Costa / Abstract: Retroviral integrase can efficiently utilise nucleosomes for insertion of the reverse-transcribed viral DNA. In face of the structural constraints imposed by the nucleosomal structure, integrase ...Retroviral integrase can efficiently utilise nucleosomes for insertion of the reverse-transcribed viral DNA. In face of the structural constraints imposed by the nucleosomal structure, integrase gains access to the scissile phosphodiester bonds by lifting DNA off the histone octamer at the site of integration. To clarify the mechanism of DNA looping by integrase, we determined a 3.9 Å resolution structure of the prototype foamy virus intasome engaged with a nucleosome core particle. The structural data along with complementary single-molecule Förster resonance energy transfer measurements reveal twisting and sliding of the nucleosomal DNA arm proximal to the integration site. Sliding the nucleosomal DNA by approximately two base pairs along the histone octamer accommodates the necessary DNA lifting from the histone H2A-H2B subunits to allow engagement with the intasome. Thus, retroviral integration into nucleosomes involves the looping-and-sliding mechanism for nucleosomal DNA repositioning, bearing unexpected similarities to chromatin remodelers. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 6rny.cif.gz | 485.2 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb6rny.ent.gz | 360.2 KB | Display | PDB format |
| PDBx/mmJSON format | 6rny.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/rn/6rnyftp://data.pdbj.org/pub/pdb/validation_reports/rn/6rny | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  4960MC  4692C  4693C  6r0cC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |



-Links
 PDBj
PDBj










































- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 5 types, 12 molecules AEBFCGDHKLOP
| #1: Protein | Mass: 15360.983 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: H3F3A, H3.3A, H3F3, PP781, H3F3B, H3.3B / Production host:  #2: Protein | Mass: 11394.426 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, ...Gene: HIST1H4A, H4/A, H4FA, HIST1H4B, H4/I, H4FI, HIST1H4C, H4/G, H4FG, HIST1H4D, H4/B, H4FB, HIST1H4E, H4/J, H4FJ, HIST1H4F, H4/C, H4FC, HIST1H4H, H4/H, H4FH, HIST1H4I, H4/M, H4FM, HIST1H4J, H4/E, H4FE, HIST1H4K, H4/D, H4FD, HIST1H4L, H4/K, H4FK, HIST2H4A, H4/N, H4F2, H4FN, HIST2H4, HIST2H4B, H4/O, H4FO, HIST4H4 Production host: #3: Protein | Mass: 14121.537 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: HIST1H2AG, H2AFP, HIST1H2AI, H2AFC, HIST1H2AK, H2AFD, HIST1H2AL, H2AFI, HIST1H2AM, H2AFN Production host: #4: Protein | Mass: 13937.213 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human)Gene: HIST1H2BC, H2BFL, HIST1H2BE, H2BFH, HIST1H2BF, H2BFG, HIST1H2BG, H2BFA, HIST1H2BI, H2BFK Production host: #5: Protein | Mass: 44456.695 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Human spumaretrovirus / Gene: pol / Production host: References: UniProt: P14350, RNA-directed DNA polymerase, DNA-directed DNA polymerase, ribonuclease H, Hydrolases; Acting on peptide bonds (peptidases); Aspartic endopeptidases, Transferases; ...References: UniProt: P14350, RNA-directed DNA polymerase, DNA-directed DNA polymerase, ribonuclease H, Hydrolases; Acting on peptide bonds (peptidases); Aspartic endopeptidases, Transferases; Transferring phosphorus-containing groups; Nucleotidyltransferases, Hydrolases; Acting on ester bonds |
|---|
-DNA chain , 5 types, 6 molecules IUTJQM
| #6: DNA chain | Mass: 39189.094 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
|---|---|
| #7: DNA chain | Mass: 33581.453 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
| #8: DNA chain | Mass: 10114.523 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
| #9: DNA chain | Mass: 16397.531 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) Homo sapiens (human) |
| #10: DNA chain | Mass: 5834.794 Da / Num. of mol.: 2 / Source method: obtained synthetically / Source: (synth.) Pyrobaculum filamentous virus 1 |
-Non-polymers , 1 types, 2 molecules 
| #11: Chemical |  Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: Mg |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
| ||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.399 MDa / Experimental value: NO | ||||||||||||||||||||||||||||||||||||||||||
| Source (natural) |
| ||||||||||||||||||||||||||||||||||||||||||
| Source (recombinant) |
| ||||||||||||||||||||||||||||||||||||||||||
| Buffer solution | pH: 7 | ||||||||||||||||||||||||||||||||||||||||||
| Buffer component | Conc.: 1 MM / Name: DTT | ||||||||||||||||||||||||||||||||||||||||||
| Specimen | Conc.: 0.1 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||||||||||||||||||||
| Specimen support | Grid material: COPPER / Grid mesh size: 400 divisions/in. / Grid type: C-flat-1/1 | ||||||||||||||||||||||||||||||||||||||||||
| Vitrification | Instrument: LEICA EM CPC / Cryogen name: ETHANE / Humidity: 80 % / Chamber temperature: 277 K / Details: 1 min incubation 3.5s blot |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 59000 X / Nominal defocus max: 3.5 nm / Nominal defocus min: 1.5 nm / Cs: 2.7 mm / C2 aperture diameter: 70 µm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Average exposure time: 1.6 sec. / Electron dose: 56 e/Å2 / Detector mode: INTEGRATING / Film or detector model: FEI FALCON II (4k x 4k) / Num. of grids imaged: 2 / Num. of real images: 4916 |
| EM imaging optics | Spherical aberration corrector: Microscope was modified with a Cs corrector (NeCEN netherlands) |
| Image scans | Movie frames/image: 7 |
- Processing
Processing
| Software | Name: PHENIX / Version: 1.14_3235: / Classification: refinement | ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EM software |
| ||||||||||||||||||||||||||||||||||||||||
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 3125 | ||||||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C1 (asymmetric) | ||||||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.9 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 177155 / Algorithm: FOURIER SPACE / Num. of class averages: 3 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||||||
| Atomic model building | Space: REAL Details: The initial model was placed in the density using Chimera. Manual building was performed in Coot and final refinement was carried out using phenix.real_space_refine and namdinator. ...Details: The initial model was placed in the density using Chimera. Manual building was performed in Coot and final refinement was carried out using phenix.real_space_refine and namdinator. Additional restraints describing protein secondary structure, DNA base pairing and stacking were used in Phenix. | ||||||||||||||||||||||||||||||||||||||||
| Atomic model building |
|