ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5iqr | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
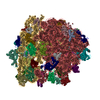
| タイトル | Structure of RelA bound to the 70S ribosome | ||||||||||||
 要素 要素 |
| ||||||||||||
 キーワード キーワード | RIBOSOME / tRNA / RelA / ppGpp | ||||||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報guanosine tetraphosphate metabolic process / guanosine-3',5'-bis(diphosphate) 3'-diphosphatase activity / GTP diphosphokinase / GTP diphosphokinase activity / guanosine tetraphosphate biosynthetic process / nucleobase-containing small molecule interconversion / negative regulation of cytoplasmic translational initiation / response to starvation / stringent response / ornithine decarboxylase inhibitor activity ...guanosine tetraphosphate metabolic process / guanosine-3',5'-bis(diphosphate) 3'-diphosphatase activity / GTP diphosphokinase / GTP diphosphokinase activity / guanosine tetraphosphate biosynthetic process / nucleobase-containing small molecule interconversion / negative regulation of cytoplasmic translational initiation / response to starvation / stringent response / ornithine decarboxylase inhibitor activity / transcription antitermination factor activity, RNA binding / misfolded RNA binding / Group I intron splicing / RNA folding / transcriptional attenuation / endoribonuclease inhibitor activity / positive regulation of ribosome biogenesis / RNA-binding transcription regulator activity / translational termination / negative regulation of cytoplasmic translation / four-way junction DNA binding / DnaA-L2 complex / translation repressor activity / negative regulation of translational initiation / regulation of mRNA stability / negative regulation of DNA-templated DNA replication initiation / mRNA regulatory element binding translation repressor activity / assembly of large subunit precursor of preribosome / positive regulation of RNA splicing / ribosome assembly / transcription elongation factor complex / regulation of DNA-templated transcription elongation / cytosolic ribosome assembly / response to reactive oxygen species / DNA endonuclease activity / transcription antitermination / translational initiation / regulation of cell growth / DNA-templated transcription termination / response to radiation / maintenance of translational fidelity / mRNA 5'-UTR binding / kinase activity / ribosome biogenesis / large ribosomal subunit / regulation of translation / transferase activity / ribosome binding / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / 5S rRNA binding / ribosomal large subunit assembly / cytosolic small ribosomal subunit / large ribosomal subunit rRNA binding / cytosolic large ribosomal subunit / cytoplasmic translation / tRNA binding / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / response to antibiotic / negative regulation of DNA-templated transcription / mRNA binding / GTP binding / DNA binding / RNA binding / zinc ion binding / ATP binding / membrane / cytosol / cytoplasm 類似検索 - 分子機能 | ||||||||||||
| 生物種 |  synthetic construct (人工物) | ||||||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3 Å | ||||||||||||
 データ登録者 データ登録者 | Brown, A. / Fernandez, I.S. / Gordiyenko, Y. / Ramakrishnan, V. | ||||||||||||
 引用 引用 | ジャーナル: Nature / 年: 2016 タイトル: Ribosome-dependent activation of stringent control. 著者: Alan Brown / Israel S Fernández / Yuliya Gordiyenko / V Ramakrishnan /  要旨: In order to survive, bacteria continually sense, and respond to, environmental fluctuations. Stringent control represents a key bacterial stress response to nutrient starvation that leads to rapid ...In order to survive, bacteria continually sense, and respond to, environmental fluctuations. Stringent control represents a key bacterial stress response to nutrient starvation that leads to rapid and comprehensive reprogramming of metabolic and transcriptional patterns. In general, transcription of genes for growth and proliferation is downregulated, while those important for survival and virulence are upregulated. Amino acid starvation is sensed by depletion of the aminoacylated tRNA pools, and this results in accumulation of ribosomes stalled with non-aminoacylated (uncharged) tRNA in the ribosomal A site. RelA is recruited to stalled ribosomes and activated to synthesize a hyperphosphorylated guanosine analogue, (p)ppGpp, which acts as a pleiotropic secondary messenger. However, structural information about how RelA recognizes stalled ribosomes and discriminates against aminoacylated tRNAs is missing. Here we present the cryo-electron microscopy structure of RelA bound to the bacterial ribosome stalled with uncharged tRNA. The structure reveals that RelA utilizes a distinct binding site compared to the translational factors, with a multi-domain architecture that wraps around a highly distorted A-site tRNA. The TGS (ThrRS, GTPase and SpoT) domain of RelA binds the CCA tail to orient the free 3' hydroxyl group of the terminal adenosine towards a β-strand, such that an aminoacylated tRNA at this position would be sterically precluded. The structure supports a model in which association of RelA with the ribosome suppresses auto-inhibition to activate synthesis of (p)ppGpp and initiate the stringent response. Since stringent control is responsible for the survival of pathogenic bacteria under stress conditions, and contributes to chronic infections and antibiotic tolerance, RelA represents a good target for the development of novel antibacterial therapeutics. | ||||||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5iqr.cif.gz | 3.8 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5iqr.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 5iqr.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 5iqr_validation.pdf.gz | 1.6 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 5iqr_full_validation.pdf.gz | 1.7 MB | 表示 | |
| XML形式データ | 5iqr_validation.xml.gz | 231.9 KB | 表示 | |
| CIF形式データ | 5iqr_validation.cif.gz | 402.9 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/iq/5iqrftp://data.pdbj.org/pub/pdb/validation_reports/iq/5iqr | HTTPS FTP |
-関連構造データ
| 関連構造データ | 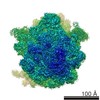 8107MC 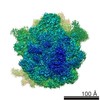 8108C 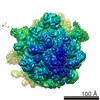 8109C  8110C  8111C  8112C  8113C 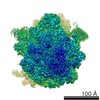 8114C 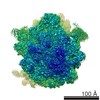 8115C M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |
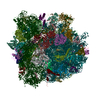
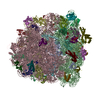
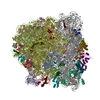
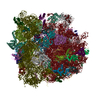
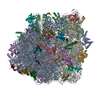
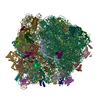
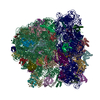
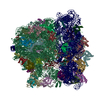
-リンク
 PDBj
PDBj































- 集合体
集合体
| 登録構造単位 | 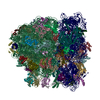
|
|---|---|
| 1 |
|
-要素
+50S ribosomal protein ... , 31種, 31分子 BCDEFGHIJKLMNOPQRSTUVWXYZabcdef
-30S ribosomal protein ... , 20種, 20分子 ghijklmnopqrstuvwxyz
| #32: タンパク質 | 分子量: 26781.670 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #33: タンパク質 | 分子量: 25975.209 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #34: タンパク質 | 分子量: 23514.199 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #35: タンパク質 | 分子量: 17629.398 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #36: タンパク質 | 分子量: 15727.512 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #37: タンパク質 | 分子量: 20055.156 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #38: タンパク質 | 分子量: 14146.557 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #39: タンパク質 | 分子量: 14886.270 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #40: タンパク質 | 分子量: 11755.597 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #41: タンパク質 | 分子量: 13870.975 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #42: タンパク質 | 分子量: 13814.249 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #43: タンパク質 | 分子量: 13128.467 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #44: タンパク質 | 分子量: 11606.560 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #45: タンパク質 | 分子量: 10290.816 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #46: タンパク質 | 分子量: 9207.572 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #47: タンパク質 | 分子量: 9724.491 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #48: タンパク質 | 分子量: 9063.508 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #49: タンパク質 | 分子量: 10455.355 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #50: タンパク質 | 分子量: 9708.464 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #51: タンパク質 | 分子量: 8524.039 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-RNA鎖 , 7種, 7分子 1234567
| #52: RNA鎖 | 分子量: 941832.625 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #53: RNA鎖 | 分子量: 497075.812 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #54: RNA鎖 | 分子量: 38177.762 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #55: RNA鎖 | 分子量: 24485.539 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #56: RNA鎖 | 分子量: 24893.010 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #57: RNA鎖 | 分子量: 24707.926 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #58: RNA鎖 | 分子量: 3146.916 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
-タンパク質 , 1種, 1分子 8
| #59: タンパク質 | 分子量: 83996.617 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|
-非ポリマー , 5種, 306分子 


| #60: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 299 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 299 / 由来タイプ: 合成 / 式: Mg#61: 化合物 |  分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 3 / 由来タイプ: 合成 / 式: Zn#62: 化合物 | ChemComp-PAR / |  分子量: 615.628 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C23H45N5O14 / コメント: 抗菌剤, 薬剤*YM 分子量: 615.628 Da / 分子数: 1 / 由来タイプ: 合成 / 式: C23H45N5O14 / コメント: 抗菌剤, 薬剤*YM#63: 化合物 | ChemComp-MET / |  タイプ: L-peptide linking / 分子量: 149.211 Da / 分子数: 1 / 由来タイプ: 天然 / 式: C5H11NO2S タイプ: L-peptide linking / 分子量: 149.211 Da / 分子数: 1 / 由来タイプ: 天然 / 式: C5H11NO2S#64: 水 | ChemComp-HOH / | 分子量: 18.015 Da / 分子数: 2 / 由来タイプ: 天然 / 式: H2O |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: RIBOSOME / タイプ: RIBOSOME / Entity ID: #1-#59 / 由来: NATURAL | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 由来(天然) | 生物種: | ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||||||||
| 緩衝液成分 |
| ||||||||||||||||||||||||||||||
| 試料 | 濃度: 0.1 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES / 詳細: This sample was monodisperse. | ||||||||||||||||||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 400 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | ||||||||||||||||||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK II / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277 K / 詳細: Grids were blotted for 5 s |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI POLARA 300 |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(補正後): 104478 X / 最大 デフォーカス(公称値): 3000 nm / 最小 デフォーカス(公称値): 1800 nm / アライメント法: COMA FREE |
| 試料ホルダ | 凍結剤: NITROGEN |
| 撮影 | 電子線照射量: 35 e/Å2 / フィルム・検出器のモデル: OTHER 詳細: FEI Falcon III Images were collected in movie-mode at 30 frames per second |
- 解析
解析
| ソフトウェア | 名称: REFMAC / バージョン: 5.8.0124 / 分類: 精密化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EMソフトウェア |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CTF補正 | タイプ: NONE | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 183615 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 98498 / 対称性のタイプ: POINT | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | B value: 112.7 / プロトコル: OTHER / 空間: RECIPROCAL / Target criteria: Average FSC | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | PDB-ID: 4YBB Accession code: 4YBB / Source name: PDB / タイプ: experimental model | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 解像度: 3→257.28 Å / Cor.coef. Fo:Fc: 0.907 / SU B: 16.623 / SU ML: 0.301 / ESU R: 0.472 立体化学のターゲット値: MAXIMUM LIKELIHOOD WITH PHASES 詳細: HYDROGENS HAVE BEEN ADDED IN THE RIDING POSITIONS
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 溶媒の処理 | 溶媒モデル: PARAMETERS FOR MASK CACLULATION | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 106.009 Å2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化ステップ | サイクル: 1 / 合計: 151800 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
|