deactivation of mitotic spindle assembly checkpoint / mitotic spindle assembly checkpoint MAD1-MAD2 complex / metaphase/anaphase transition of cell cycle / metaphase/anaphase transition of meiosis I / Inhibition of the proteolytic activity of APC/C required for the onset of anaphase by mitotic spindle checkpoint components / mitotic checkpoint complex / positive regulation of ubiquitin protein ligase activity / meiotic recombination checkpoint signaling / negative regulation of ubiquitin protein ligase activity / positive regulation of anaphase-promoting complex-dependent catabolic process ...deactivation of mitotic spindle assembly checkpoint / mitotic spindle assembly checkpoint MAD1-MAD2 complex / metaphase/anaphase transition of cell cycle / metaphase/anaphase transition of meiosis I / Inhibition of the proteolytic activity of APC/C required for the onset of anaphase by mitotic spindle checkpoint components / mitotic checkpoint complex / positive regulation of ubiquitin protein ligase activity / meiotic recombination checkpoint signaling / negative regulation of ubiquitin protein ligase activity / positive regulation of anaphase-promoting complex-dependent catabolic process / positive regulation of synapse maturation / regulation of meiotic nuclear division / Conversion from APC/C:Cdc20 to APC/C:Cdh1 in late anaphase / Inactivation of APC/C via direct inhibition of the APC/C complex / APC/C:Cdc20 mediated degradation of mitotic proteins / positive regulation of mitotic cell cycle spindle assembly checkpoint / regulation of dendrite development / synaptonemal complex assembly / anaphase-promoting complex / Phosphorylation of Emi1 / anaphase-promoting complex-dependent catabolic process / regulation of meiotic cell cycle / anaphase-promoting complex binding / regulation of exit from mitosis / nuclear pore nuclear basket / positive regulation of synaptic plasticity / positive regulation of mitotic metaphase/anaphase transition / negative regulation of mitotic cell cycle / oocyte maturation / ubiquitin ligase activator activity / female meiosis I / reciprocal meiotic recombination / mitotic sister chromatid cohesion / mitotic spindle assembly checkpoint signaling / oogenesis / Regulation of APC/C activators between G1/S and early anaphase / mitotic sister chromatid segregation / male meiosis I / spermatid development / mitotic spindle assembly / ubiquitin-like ligase-substrate adaptor activity / Amplification of signal from unattached kinetochores via a MAD2 inhibitory signal / APC/C:Cdc20 mediated degradation of Cyclin B / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / regulation of mitotic cell cycle / APC-Cdc20 mediated degradation of Nek2A / Resolution of Sister Chromatid Cohesion / male germ cell nucleus / APC/C:Cdc20 mediated degradation of Securin / transcription coregulator activity / SCF-beta-TrCP mediated degradation of Emi1 / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / RHO GTPases Activate Formins / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / negative regulation of protein catabolic process / kinetochore / spindle / histone deacetylase binding / spindle pole / mitotic spindle / Separation of Sister Chromatids / nervous system development / double-strand break repair / Antigen processing: Ubiquitination & Proteasome degradation / chromosome / spermatogenesis / nuclear membrane / transcription by RNA polymerase II / cell differentiation / Ub-specific processing proteases / protein ubiquitination / cell division / positive regulation of cell population proliferation / centrosome / nucleolus / perinuclear region of cytoplasm / protein homodimerization activity / ATP hydrolysis activity / nucleoplasm / ATP binding / identical protein binding / nucleus / cytoplasm / cytosol 類似検索 - 分子機能
Mad1/Cdc20-bound-Mad2 binding protein / : / Mad1 and Cdc20-bound-Mad2 binding / : / TRIP13-like, AAA+ ATPase lid C-terminal domain / TRIP13 N-terminal domain / Pachytene checkpoint protein 2-like / : / CDC20/Fizzy WD40 domain / The WD repeat Cdc20/Fizzy family ...Mad1/Cdc20-bound-Mad2 binding protein / : / Mad1 and Cdc20-bound-Mad2 binding / : / TRIP13-like, AAA+ ATPase lid C-terminal domain / TRIP13 N-terminal domain / Pachytene checkpoint protein 2-like / : / CDC20/Fizzy WD40 domain / The WD repeat Cdc20/Fizzy family / Mad2-like / HORMA domain / HORMA domain / HORMA domain profile. / HORMA domain superfamily / ClpA/B family / ATPase, AAA-type, conserved site / AAA-protein family signature. / ATPase family associated with various cellular activities (AAA) / ATPase, AAA-type, core / Trp-Asp (WD) repeats signature. / Trp-Asp (WD) repeats profile. / Trp-Asp (WD) repeats circular profile. / WD40 repeats / WD40 repeat / WD40-repeat-containing domain superfamily / WD40/YVTN repeat-like-containing domain superfamily / ATPases associated with a variety of cellular activities / AAA+ ATPase domain / P-loop containing nucleoside triphosphate hydrolase 類似検索 - ドメイン・相同性
Cell division cycle protein 20 homolog / Mitotic spindle assembly checkpoint protein MAD2A / MAD2L1-binding protein / Pachytene checkpoint protein 2 homolog 類似検索 - 構成要素
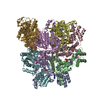
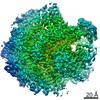
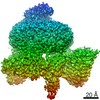
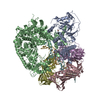
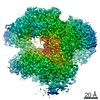
ジャーナル: Nature / 年: 2018 タイトル: Mechanism for remodelling of the cell cycle checkpoint protein MAD2 by the ATPase TRIP13. 著者: Claudio Alfieri / Leifu Chang / David Barford / 要旨: The maintenance of genome stability during mitosis is coordinated by the spindle assembly checkpoint (SAC) through its effector the mitotic checkpoint complex (MCC), an inhibitor of the anaphase- ...The maintenance of genome stability during mitosis is coordinated by the spindle assembly checkpoint (SAC) through its effector the mitotic checkpoint complex (MCC), an inhibitor of the anaphase-promoting complex (APC/C, also known as the cyclosome). Unattached kinetochores control MCC assembly by catalysing a change in the topology of the β-sheet of MAD2 (an MCC subunit), thereby generating the active closed MAD2 (C-MAD2) conformer. Disassembly of free MCC, which is required for SAC inactivation and chromosome segregation, is an ATP-dependent process driven by the AAA+ ATPase TRIP13. In combination with p31, an SAC antagonist, TRIP13 remodels C-MAD2 into inactive open MAD2 (O-MAD2). Here, we present a mechanism that explains how TRIP13-p31 disassembles the MCC. Cryo-electron microscopy structures of the TRIP13-p31-C-MAD2-CDC20 complex reveal that p31 recruits C-MAD2 to a defined site on the TRIP13 hexameric ring, positioning the N terminus of C-MAD2 (MAD2) to insert into the axial pore of TRIP13 and distorting the TRIP13 ring to initiate remodelling. Molecular modelling suggests that by gripping MAD2 within its axial pore, TRIP13 couples sequential ATP-driven translocation of its hexameric ring along MAD2 to push upwards on, and simultaneously rotate, the globular domains of the p31-C-MAD2 complex. This unwinds a region of the αA helix of C-MAD2 that is required to stabilize the C-MAD2 β-sheet, thus destabilizing C-MAD2 in favour of O-MAD2 and dissociating MAD2 from p31. Our study provides insights into how specific substrates are recruited to AAA+ ATPases through adaptor proteins and suggests a model of how translocation through the axial pore of AAA+ ATPases is coupled to protein remodelling.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 英国, 1件
英国, 1件  引用
引用
 構造の表示
構造の表示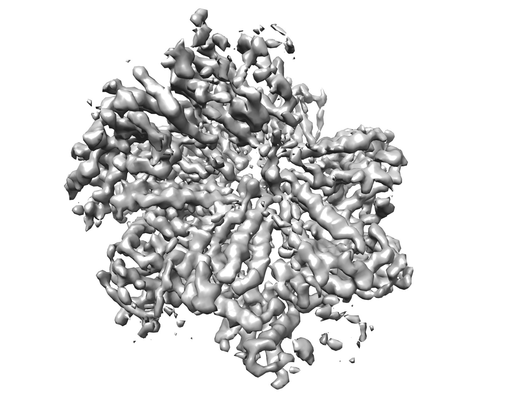
 ダウンロードとリンク
ダウンロードとリンク emd_4166.png
emd_4166.png http://ftp.pdbj.org/pub/emdb/structures/EMD-4166
http://ftp.pdbj.org/pub/emdb/structures/EMD-4166














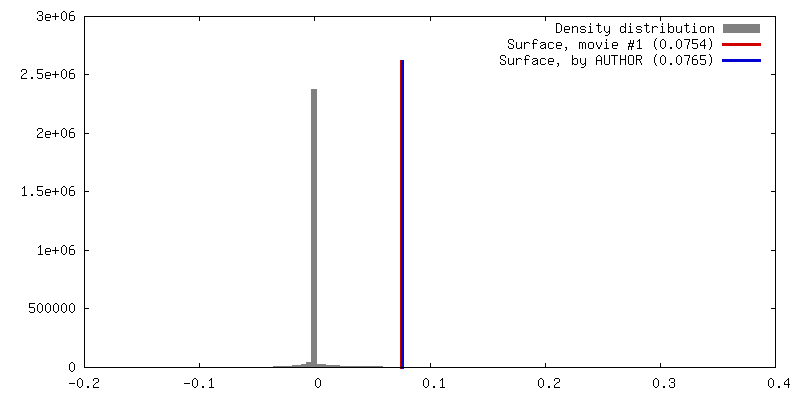
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)

























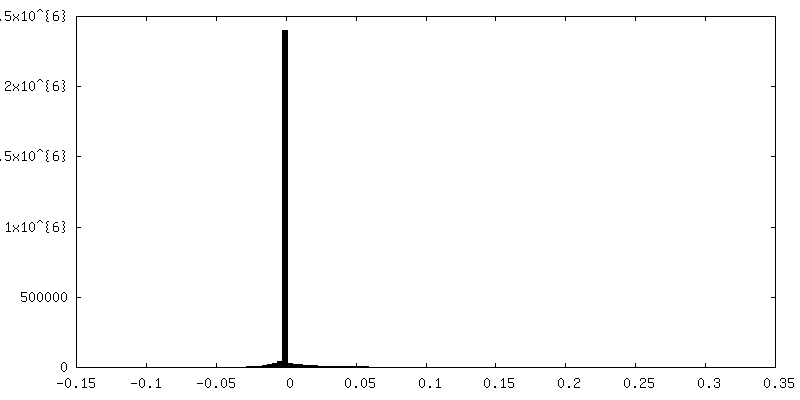
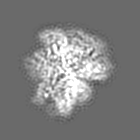

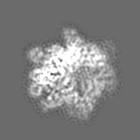

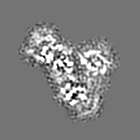

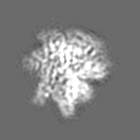



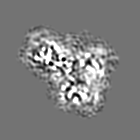



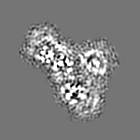
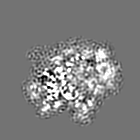





 試料の構成要素
試料の構成要素
 Trichoplusia ni (イラクサキンウワバ)
Trichoplusia ni (イラクサキンウワバ)
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN
