 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-2796 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | Membrane bound pleurotolysin prepore (TMH2 strand lock) trapped with engineered disulphide cross-link | |||||||||
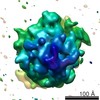
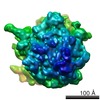
 マップデータ マップデータ | This TMH2 strand disulphide trap mutant (V277C, K291C) was engineered on a background PlyB variant that lacks the wildtype cysteine (C487A) in order to avoid incorrect disulphide bond formation. | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | MACPF/CDC superfamily / pore-forming proteins | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報 | |||||||||
| 生物種 |  Pleurotus ostreatus (ヒラタケ) Pleurotus ostreatus (ヒラタケ) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 14.0 Å | |||||||||
 データ登録者 データ登録者 | Lukoyanova N / Kondos SC / Farabella I / Law RHP / Reboul CF / Caradoc-Davies TT / Spicer BA / Kleifeld O / Perugini M / Ekkel S ...Lukoyanova N / Kondos SC / Farabella I / Law RHP / Reboul CF / Caradoc-Davies TT / Spicer BA / Kleifeld O / Perugini M / Ekkel S / Hatfaludi T / Oliver K / Hotze EM / Tweten RK / Whisstock JC / Topf M / Dunstone MA / Saibil HR | |||||||||
 引用 引用 | ジャーナル: PLoS Biol / 年: 2015 タイトル: Conformational changes during pore formation by the perforin-related protein pleurotolysin. 著者: Natalya Lukoyanova / Stephanie C Kondos / Irene Farabella / Ruby H P Law / Cyril F Reboul / Tom T Caradoc-Davies / Bradley A Spicer / Oded Kleifeld / Daouda A K Traore / Susan M Ekkel / Ilia ...著者: Natalya Lukoyanova / Stephanie C Kondos / Irene Farabella / Ruby H P Law / Cyril F Reboul / Tom T Caradoc-Davies / Bradley A Spicer / Oded Kleifeld / Daouda A K Traore / Susan M Ekkel / Ilia Voskoboinik / Joseph A Trapani / Tamas Hatfaludi / Katherine Oliver / Eileen M Hotze / Rodney K Tweten / James C Whisstock / Maya Topf / Helen R Saibil / Michelle A Dunstone /    要旨: Membrane attack complex/perforin-like (MACPF) proteins comprise the largest superfamily of pore-forming proteins, playing crucial roles in immunity and pathogenesis. Soluble monomers assemble into ...Membrane attack complex/perforin-like (MACPF) proteins comprise the largest superfamily of pore-forming proteins, playing crucial roles in immunity and pathogenesis. Soluble monomers assemble into large transmembrane pores via conformational transitions that remain to be structurally and mechanistically characterised. Here we present an 11 Å resolution cryo-electron microscopy (cryo-EM) structure of the two-part, fungal toxin Pleurotolysin (Ply), together with crystal structures of both components (the lipid binding PlyA protein and the pore-forming MACPF component PlyB). These data reveal a 13-fold pore 80 Å in diameter and 100 Å in height, with each subunit comprised of a PlyB molecule atop a membrane bound dimer of PlyA. The resolution of the EM map, together with biophysical and computational experiments, allowed confident assignment of subdomains in a MACPF pore assembly. The major conformational changes in PlyB are a ∼70° opening of the bent and distorted central β-sheet of the MACPF domain, accompanied by extrusion and refolding of two α-helical regions into transmembrane β-hairpins (TMH1 and TMH2). We determined the structures of three different disulphide bond-trapped prepore intermediates. Analysis of these data by molecular modelling and flexible fitting allows us to generate a potential trajectory of β-sheet unbending. The results suggest that MACPF conformational change is triggered through disruption of the interface between a conserved helix-turn-helix motif and the top of TMH2. Following their release we propose that the transmembrane regions assemble into β-hairpins via top down zippering of backbone hydrogen bonds to form the membrane-inserted β-barrel. The intermediate structures of the MACPF domain during refolding into the β-barrel pore establish a structural paradigm for the transition from soluble monomer to pore, which may be conserved across the whole superfamily. The TMH2 region is critical for the release of both TMH clusters, suggesting why this region is targeted by endogenous inhibitors of MACPF function. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |
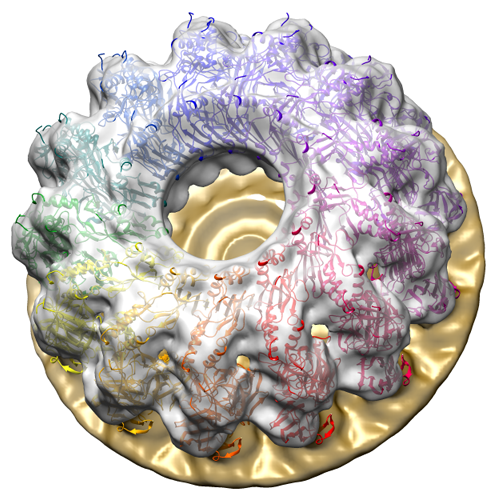
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_2796.map.gz | 28.4 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-2796-v30.xmlemd-2796.xml | 14.3 KB 14.3 KB | 表示 表示 | EMDBヘッダ |
| 画像 | 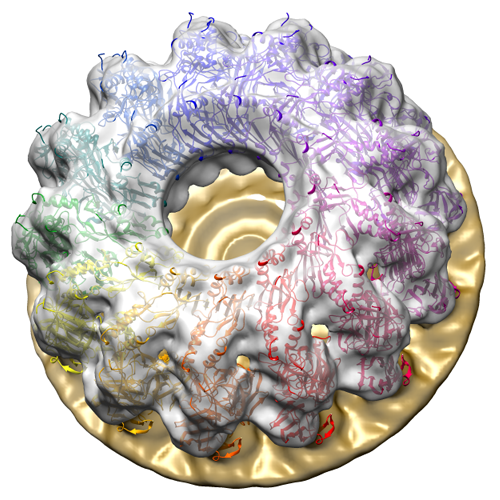 EMD-2796_image.png EMD-2796_image.png | 336.1 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-2796ftp://ftp.pdbj.org/pub/emdb/structures/EMD-2796 http://ftp.pdbj.org/pub/emdb/structures/EMD-2796ftp://ftp.pdbj.org/pub/emdb/structures/EMD-2796 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_2796_validation.pdf.gz | 210.7 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_2796_full_validation.pdf.gz | 209.8 KB | 表示 | |
| XML形式データ | emd_2796_validation.xml.gz | 5 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-2796ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-2796 | HTTPS FTP |
-関連構造データ
| 関連構造データ |  4v3nMC  2793C  2794C  2795C  4oebC  4oejC  4ov8C  4v2tC  4v3aC  4v3mC M: このマップから作成された原子モデル C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ |


-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|
-マップ
| ファイル | ダウンロード / ファイル: emd_2796.map.gz / 形式: CCP4 / 大きさ: 29.8 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | This TMH2 strand disulphide trap mutant (V277C, K291C) was engineered on a background PlyB variant that lacks the wildtype cysteine (C487A) in order to avoid incorrect disulphide bond formation. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 2 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
- 試料の構成要素
試料の構成要素
-全体 : pleurotolysin prepore on liposomes (TMH2 strand lock) trapped wit...
| 全体 | 名称: pleurotolysin prepore on liposomes (TMH2 strand lock) trapped with engineered disulphide |
|---|---|
| 要素 |
|
-超分子 #1000: pleurotolysin prepore on liposomes (TMH2 strand lock) trapped wit...
| 超分子 | 名称: pleurotolysin prepore on liposomes (TMH2 strand lock) trapped with engineered disulphide タイプ: sample / ID: 1000 詳細: 13 fold symmetrical ring oligomers of 26 PlyA and 13 PlyB (C487A, V277C, K291C variant) molecules on liposomes Number unique components: 2 |
|---|
-分子 #1: pleurotolysin A
| 分子 | 名称: pleurotolysin A / タイプ: protein_or_peptide / ID: 1 / Name.synonym: PlyA / コピー数: 26 / 組換発現: Yes |
|---|---|
| 由来(天然) | 生物種: Pleurotus ostreatus (ヒラタケ) / 別称: Oyster mushroom, White-rot fungus |
| 分子量 | 理論値: 17 KDa |
| 組換発現 | 生物種:  |
| 配列 | UniProtKB: Pleurotolysin A / GO: hemolysis by symbiont of host erythrocytes / InterPro: Hemolysin, aegerolysin type |
-分子 #2: pleurotolysin B
| 分子 | 名称: pleurotolysin B / タイプ: protein_or_peptide / ID: 2 / Name.synonym: PlyB / 詳細: C487A, V277C, K291C variant / コピー数: 13 / 組換発現: Yes |
|---|---|
| 由来(天然) | 生物種: Pleurotus ostreatus (ヒラタケ) / 別称: Oyster mushroom, White-rot fungus |
| 分子量 | 理論値: 52 KDa |
| 組換発現 | 生物種: |
| 配列 | UniProtKB: Pleurotolysin B / GO: hemolysis by symbiont of host erythrocytes InterPro: Membrane attack complex component/perforin (MACPF) domain |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 濃度 | 0.02 mg/mL |
|---|---|
| 緩衝液 | pH: 7.4 / 詳細: 50 mM NaCl, 20 mM Hepes |
| グリッド | 詳細: 300 mesh lacey copper grids |
| 凍結 | 凍結剤: ETHANE / チャンバー内湿度: 80 % / 装置: FEI VITROBOT MARK III 手法: Pleurotolysin A was first added to sphingomyelin/cholesterol liposomes at a molar ratio of 1:2000 protein to lipid in the above buffer. After 5 min incubation at room temperature, ...手法: Pleurotolysin A was first added to sphingomyelin/cholesterol liposomes at a molar ratio of 1:2000 protein to lipid in the above buffer. After 5 min incubation at room temperature, pleurotolysin B was added to the mixture at a molar ratio of 1:2 to pleurotolysin A. The mixture was incubated at 40 C or room temperature for 30 min after which 3.5 uL were placed on negatively glow discharged lacey grids and vitrified in liquid ethane using a Vitrobot. Blotting was carried out at 36 C and 80% humidity. |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI POLARA 300 |
|---|---|
| 温度 | 最低: 90 K / 最高: 102 K / 平均: 94 K |
| アライメント法 | Legacy - 非点収差: Objective lens astigmatism was corrected at 115,000 times magnification |
| 日付 | 2013年3月5日 |
| 撮影 | カテゴリ: CCD フィルム・検出器のモデル: GATAN ULTRASCAN 4000 (4k x 4k) デジタル化 - サンプリング間隔: 15 µm / 実像数: 210 / 平均電子線量: 25 e/Å2 / ビット/ピクセル: 16 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 倍率(補正後): 76148 / 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / Cs: 2.3 mm / 最大 デフォーカス(公称値): 3.4 µm / 最小 デフォーカス(公称値): 0.8 µm / 倍率(公称値): 59000 |
| 試料ステージ | 試料ホルダー: liquid nitrogen cooled / 試料ホルダーモデル: GATAN HELIUM |
| 実験機器 |  モデル: Tecnai Polara / 画像提供: FEI Company |
-画像解析
| 詳細 | 2,700 prepore side views with adjacent membrane regions were extracted from liposome images and treated as single particles. These particles were initially used to separate 12- and 13-fold symmetries by angular reconstitution followed by competitive projection matching. |
|---|---|
| CTF補正 | 詳細: Estimated with CTFFIND3, then phases flipped for each particle |
| 最終 再構成 | 想定した対称性 - 点群: C13 (13回回転対称) / アルゴリズム: OTHER / 解像度のタイプ: BY AUTHOR / 解像度: 14.0 Å / 解像度の算出法: OTHER / ソフトウェア - 名称: Imagic, Spider / 詳細: SPIDER operation BP RP was used for reconstruction / 使用した粒子像数: 1110 |
-原子モデル構築 1
| 初期モデル | PDB ID: |
|---|---|
| ソフトウェア | 名称: Chimera |
| 精密化 | 空間: REAL / プロトコル: RIGID BODY FIT 当てはまり具合の基準: cross-correlation coefficient |
| 得られたモデル | PDB-4v3n: |
