 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-4oeb: Structure of membrane binding protein pleurotolysin A from Pleuro... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4oeb | ||||||
|---|---|---|---|---|---|---|---|
| Title | Structure of membrane binding protein pleurotolysin A from Pleurotus ostreatus | ||||||
 Components Components | Pleurotolysin A | ||||||
 Keywords Keywords | MEMBRANE BINDING PROTEIN / beta-sandwich fold / pleurotolysin B | ||||||
| Function / homology | Mutm (Fpg) Protein; Chain: A, domain 2 - #50 / Hemolysin, aegerolysin type / Aegerolysin / Mutm (Fpg) Protein; Chain: A, domain 2 / symbiont-mediated hemolysis of host erythrocyte / Sandwich / Mainly Beta / Pleurotolysin A Function and homology information Function and homology information | ||||||
| Biological species |  Pleurotus ostreatus (oyster mushroom) Pleurotus ostreatus (oyster mushroom) | ||||||
| Method |  X-RAY DIFFRACTION / SYNCHROTRON / SAD / Resolution: 1.85 Å X-RAY DIFFRACTION / SYNCHROTRON / SAD / Resolution: 1.85 Å | ||||||
 Authors Authors | Dunstone, M.A. / Caradoc-Davies, T.T. / Whisstock, J.C. / Law, R.H.P. | ||||||
 Citation Citation | Journal: PLoS Biol / Year: 2015 Title: Conformational changes during pore formation by the perforin-related protein pleurotolysin. Authors: Natalya Lukoyanova / Stephanie C Kondos / Irene Farabella / Ruby H P Law / Cyril F Reboul / Tom T Caradoc-Davies / Bradley A Spicer / Oded Kleifeld / Daouda A K Traore / Susan M Ekkel / Ilia ...Authors: Natalya Lukoyanova / Stephanie C Kondos / Irene Farabella / Ruby H P Law / Cyril F Reboul / Tom T Caradoc-Davies / Bradley A Spicer / Oded Kleifeld / Daouda A K Traore / Susan M Ekkel / Ilia Voskoboinik / Joseph A Trapani / Tamas Hatfaludi / Katherine Oliver / Eileen M Hotze / Rodney K Tweten / James C Whisstock / Maya Topf / Helen R Saibil / Michelle A Dunstone /    Abstract: Membrane attack complex/perforin-like (MACPF) proteins comprise the largest superfamily of pore-forming proteins, playing crucial roles in immunity and pathogenesis. Soluble monomers assemble into ...Membrane attack complex/perforin-like (MACPF) proteins comprise the largest superfamily of pore-forming proteins, playing crucial roles in immunity and pathogenesis. Soluble monomers assemble into large transmembrane pores via conformational transitions that remain to be structurally and mechanistically characterised. Here we present an 11 Å resolution cryo-electron microscopy (cryo-EM) structure of the two-part, fungal toxin Pleurotolysin (Ply), together with crystal structures of both components (the lipid binding PlyA protein and the pore-forming MACPF component PlyB). These data reveal a 13-fold pore 80 Å in diameter and 100 Å in height, with each subunit comprised of a PlyB molecule atop a membrane bound dimer of PlyA. The resolution of the EM map, together with biophysical and computational experiments, allowed confident assignment of subdomains in a MACPF pore assembly. The major conformational changes in PlyB are a ∼70° opening of the bent and distorted central β-sheet of the MACPF domain, accompanied by extrusion and refolding of two α-helical regions into transmembrane β-hairpins (TMH1 and TMH2). We determined the structures of three different disulphide bond-trapped prepore intermediates. Analysis of these data by molecular modelling and flexible fitting allows us to generate a potential trajectory of β-sheet unbending. The results suggest that MACPF conformational change is triggered through disruption of the interface between a conserved helix-turn-helix motif and the top of TMH2. Following their release we propose that the transmembrane regions assemble into β-hairpins via top down zippering of backbone hydrogen bonds to form the membrane-inserted β-barrel. The intermediate structures of the MACPF domain during refolding into the β-barrel pore establish a structural paradigm for the transition from soluble monomer to pore, which may be conserved across the whole superfamily. The TMH2 region is critical for the release of both TMH clusters, suggesting why this region is targeted by endogenous inhibitors of MACPF function. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4oeb.cif.gz | 229.9 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4oeb.ent.gz | 186.9 KB | Display | PDB format |
| PDBx/mmJSON format | 4oeb.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/oe/4oebftp://data.pdbj.org/pub/pdb/validation_reports/oe/4oeb | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  2793C  2794C  2795C  2796C  4oejC  4ov8C  4v2tC  4v3aC  4v3mC  4v3nC C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj- Assembly
Assembly
| Deposited unit | 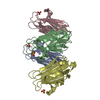
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 
| ||||||||
| 2 | 
| ||||||||
| 3 | 
| ||||||||
| 4 | 
| ||||||||
| Unit cell |
|
-Components
| #1: Protein | Mass: 15248.686 Da / Num. of mol.: 4 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Pleurotus ostreatus (oyster mushroom) / Gene: PlyA, plyA / Production host:  #2: Chemical | ChemComp-SO4 / 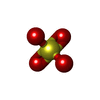  Mass: 96.063 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: SO4 Mass: 96.063 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: SO4#3: Water | ChemComp-HOH / |  Mass: 18.015 Da / Num. of mol.: 498 / Source method: isolated from a natural source / Formula: H2O Mass: 18.015 Da / Num. of mol.: 498 / Source method: isolated from a natural source / Formula: H2OHas protein modification | Y | |
|---|
-Experimental details
-Experiment
| Experiment | Method: X-RAY DIFFRACTION / Number of used crystals: 1 |
|---|
- Sample preparation
Sample preparation
| Crystal | Density Matthews: 2.68 Å3/Da / Density % sol: 54.05 % |
|---|---|
| Crystal grow | Temperature: 293 K / Method: vapor diffusion, hanging drop / pH: 6 Details: 20 mM sodium cacodylate, pH 6.0, 14% PEG3350, 0.2 M magnesium sulfate, VAPOR DIFFUSION, HANGING DROP, temperature 293K |
-Data collection
| Diffraction | Mean temperature: 100 K |
|---|---|
| Diffraction source | Source: SYNCHROTRON / Site: Australian Synchrotron / Beamline: MX1 / Wavelength: 0.95364 Å |
| Detector | Type: ADSC QUANTUM 210r / Detector: CCD / Date: Apr 16, 2008 |
| Radiation | Monochromator: double crystal Si(111) / Protocol: SINGLE WAVELENGTH / Monochromatic (M) / Laue (L): M / Scattering type: x-ray |
| Radiation wavelength | Wavelength: 0.95364 Å / Relative weight: 1 |
| Reflection | Resolution: 1.85→43.109 Å / Num. all: 54873 / Num. obs: 54873 / % possible obs: 100 % / Observed criterion σ(F): 2 / Observed criterion σ(I): 2 / Redundancy: 88.1 % / Biso Wilson estimate: 27.25 Å2 / Rmerge(I) obs: 0.113 / Net I/σ(I): 49.9 |
| Reflection shell | Resolution: 1.85→2.11 Å / Redundancy: 87.5 % / Rmerge(I) obs: 0.561 / Mean I/σ(I) obs: 11.5 / Num. unique all: 6421 / % possible all: 100 |
- Processing
Processing
| Software |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Refinement | Method to determine structure: SAD / Resolution: 1.85→43.109 Å / Cor.coef. Fo:Fc: 0.9396 / Cor.coef. Fo:Fc free: 0.9291 / SU R Cruickshank DPI: 0.115 / Cross valid method: THROUGHOUT / σ(F): 0 / SU R Blow DPI: 0.119 / SU Rfree Blow DPI: 0.109 / SU Rfree Cruickshank DPI: 0.107
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Displacement parameters | Biso mean: 35.26 Å2
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine analyze | Luzzati coordinate error obs: 0.259 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement step | Cycle: LAST / Resolution: 1.85→43.109 Å
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| LS refinement shell | Resolution: 1.85→1.9 Å / Total num. of bins used: 20
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS params. | Method: refined / Refine-ID: X-RAY DIFFRACTION
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Refinement TLS group |
|
