 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: EMDB / ID: EMD-10347 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
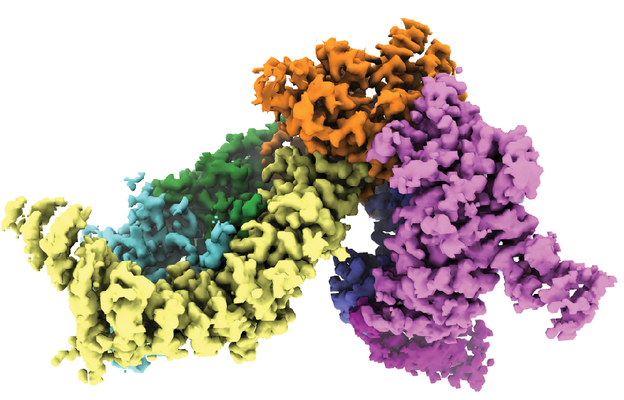
| タイトル | Structure of the SMG1-SMG8-SMG9 complex | |||||||||
 マップデータ マップデータ | CryoEM map of the SMG1-SMG8-SMG9 complex core | |||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | Kinase / NMD / IP6 / G-fold protein / PIKK family / SIGNALING PROTEIN | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報regulation of protein kinase activity / diacylglycerol-dependent serine/threonine kinase activity / chromatoid body / eye development / regulation of telomere maintenance / nuclear-transcribed mRNA catabolic process, nonsense-mediated decay / telomeric DNA binding / phosphatidylinositol phosphate biosynthetic process / mRNA export from nucleus / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) ...regulation of protein kinase activity / diacylglycerol-dependent serine/threonine kinase activity / chromatoid body / eye development / regulation of telomere maintenance / nuclear-transcribed mRNA catabolic process, nonsense-mediated decay / telomeric DNA binding / phosphatidylinositol phosphate biosynthetic process / mRNA export from nucleus / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / brain development / peptidyl-serine phosphorylation / heart development / protein autophosphorylation / in utero embryonic development / non-specific serine/threonine protein kinase / protein kinase activity / protein serine kinase activity / DNA repair / protein serine/threonine kinase activity / DNA damage response / negative regulation of apoptotic process / RNA binding / nucleoplasm / ATP binding / metal ion binding / identical protein binding / nucleus / cytosol / cytoplasm 類似検索 - 分子機能 | |||||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.45 Å | |||||||||
 データ登録者 データ登録者 | Gat Y / Schuller JM | |||||||||
| 資金援助 |  ドイツ, 1件 ドイツ, 1件
| |||||||||
 引用 引用 | ジャーナル: Nat Struct Mol Biol / 年: 2019 タイトル: InsP binding to PIKK kinases revealed by the cryo-EM structure of an SMG1-SMG8-SMG9 complex. 著者: Yair Gat / Jan Michael Schuller / Mahesh Lingaraju / Elisabeth Weyher / Fabien Bonneau / Mike Strauss / Peter J Murray / Elena Conti /  要旨: We report the 3.45-Å resolution cryo-EM structure of human SMG1-SMG8-SMG9, a phosphatidylinositol-3-kinase (PI(3)K)-related protein kinase (PIKK) complex central to messenger RNA surveillance. ...We report the 3.45-Å resolution cryo-EM structure of human SMG1-SMG8-SMG9, a phosphatidylinositol-3-kinase (PI(3)K)-related protein kinase (PIKK) complex central to messenger RNA surveillance. Structural and MS analyses reveal the presence of inositol hexaphosphate (InsP) in the SMG1 kinase. We show that the InsP-binding site is conserved in mammalian target of rapamycin (mTOR) and potentially other PIKK members, and that it is required for optimal in vitro phosphorylation of both SMG1 and mTOR substrates. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | EMマップ: SurfViewMolmilJmol/JSmol |
| 添付画像 |
- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_10347.map.gz | 3.6 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-10347-v30.xmlemd-10347.xml | 25 KB 25 KB | 表示 表示 | EMDBヘッダ |
| 画像 | 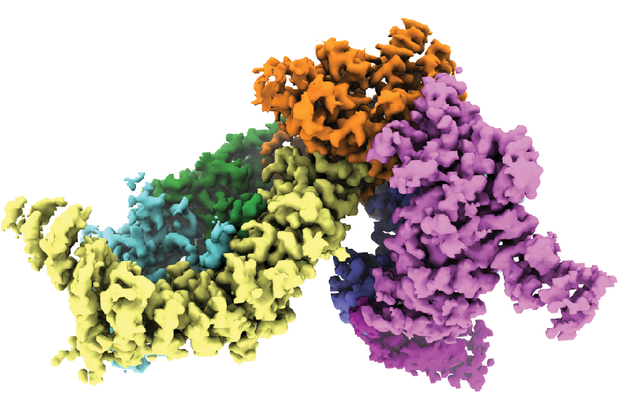 emd_10347.png emd_10347.png | 243.8 KB | ||
| Filedesc metadata | emd-10347.cif.gz | 9.1 KB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-10347ftp://ftp.pdbj.org/pub/emdb/structures/EMD-10347 http://ftp.pdbj.org/pub/emdb/structures/EMD-10347ftp://ftp.pdbj.org/pub/emdb/structures/EMD-10347 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_10347_validation.pdf.gz | 220.3 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_10347_full_validation.pdf.gz | 219.4 KB | 表示 | |
| XML形式データ | emd_10347_validation.xml.gz | 6.5 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-10347ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-10347 | HTTPS FTP |
-関連構造データ




-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |





-マップ
| ファイル | ダウンロード / ファイル: emd_10347.map.gz / 形式: CCP4 / 大きさ: 103 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 注釈 | CryoEM map of the SMG1-SMG8-SMG9 complex core | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 投影像・断面図 | 画像のコントロール
画像は Spider により作成 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ボクセルのサイズ | X=Y=Z: 1.06 Å | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 詳細 | EMDB XML:
CCP4マップ ヘッダ情報:
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















-添付データ
- 試料の構成要素
試料の構成要素
-全体 : SMG1-SMG8-SMG9 PIKK Kinase complex
| 全体 | 名称: SMG1-SMG8-SMG9 PIKK Kinase complex |
|---|---|
| 要素 |
|
-超分子 #1: SMG1-SMG8-SMG9 PIKK Kinase complex
| 超分子 | 名称: SMG1-SMG8-SMG9 PIKK Kinase complex / タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1-#3 |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
-分子 #1: SMG1,Serine/threonine-protein kinase SMG1,SMG1,Serine/threonine-p...
| 分子 | 名称: SMG1,Serine/threonine-protein kinase SMG1,SMG1,Serine/threonine-protein kinase SMG1,SMG1,Serine/threonine-protein kinase SMG1,SMG1,Serine/threonine-protein kinase SMG1,SMG1,Serine/threonine-protein kinase SMG1 タイプ: protein_or_peptide / ID: 1 詳細: Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of ...詳細: Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R,Inactive D2335A mutant of SMG1. The automatic alignment is wrong. With the numbering here - residues 2486 to the end (from the coordinates) should be aligned with residues 3659 to the end of the sequence. The following residues are modeled as Alanine 147-156 162-175 191-201 207-224 248-265 267-285 290-304 311-312 1646-1657 1663-1677 1703-1717 1962-1965 1967-1978 2006-2021 2068-2083 2248-2265 Residues 1-52 are a tag. our construct has a point mutation compared to the annotated sequence - K743R コピー数: 1 / 光学異性体: LEVO / EC番号: non-specific serine/threonine protein kinase |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 401.403938 KDa |
| 組換発現 | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) ...文字列: (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)FSTNFR DTVDILVGWH IDHTQKPSLT QQVS GWLQS LEPFWVADLA FSTTLLGQFL EDMEAYAEDL SHVASGESVD EDVPPPSVSL PKLAALLRVF STVVRSIGER FSPIR GPPI TEAYVTDVLY RVMRCVTAAN QVFFSEAVLT AANECVGVLL GSLDPSMTIH CDMVITYGLD QLENCQTCGT DYIISV LNL LTLIVEQINT KLPSSFVEKL FIPSSKLLFL RYHKEKEVVA VAHAVYQAVL SLKNIPVLET AYKLILGEMT CALNNLL HS LQLPEACSEI KHEAFKNHVF NVDNAKFVVI FDLSALTTIG NAKNSLIGMW ALSPTVFALL SKNLMIVHSD LAVHFPAI Q YAVLYTLYSH CTRHDHFISS SLSSSSPSLF DGAVISTVTT ATKKHFSIIL NLLGILLKKD NLNQDTRKLL MTWALEAAV LMRKSETYAP LFSLPSFHKF CKGLLANTLV EDVNICLQAC SSLHALSSSL PDDLLQRCVD VCRVQLVHSG TRIRQAFGKL LKSIPLDVV LSNNNHTEIQ EISLALRSHM SKAPSNTFHP QDFSDVISFI LYGNSHRTGK DNWLERLFYS CQRLDKRDQS T IPRNLLKT DAVLWQWAIW EAAQFTVLSK LRTPLGRAQD TFQTIEGIIR SLAAHTLNPD QDVSQWTTAD NDEGHGNNQL RL VLLLQYL ENLEKLMYNA YEGCANALTS PPKVIRTFFY TNRQTCQDWL TRIRLSIMRV GLLAGQPAVT VRHGFDLLTE MKT TSLSQG NELEVTIMMV VEALCELHCP EAIQGIAVWS SSIVGKNLLW INSVAQQAEG RFEKASVEYQ EHLCAMTGVD CCIS SFDKS VLTLANAGRN SASPKHSLNG ESRKTVLSKP TDSSPEVINY LGNKACECYI SIADWAAVQE WQNAIHDLKK STSST SLNL KADFNYIKSL SSFESGKFVE CTEQLELLPG ENINLLAGGS KEKIDMKKLL PNMLSPDPRE LQKSIEVQLL RSSVCL ATA LNPIEQDQKW QSITENVVKY LKQTSRIAIG PLRLSTLTVS QSLPVLSTLQ LYCSSALENT VSNRLSTEDC LIPLFSE AL RSCKQHDVRP WMQALRYTMY QNQLLEKIKE QTVPIRSHLM ELGLTAAKFA RKRGNVSLAT RLLAQCSEVQ LGKTTTAQ D LVQHFKKLST QGQVDEKWGP ELDIEKTKLL YTAGQSTHAM EMLSSCAISF CKSVKAEYAV AKSILTLAKW IQAEWKEIS GQLKQVYRAQ HQQNFTGLST LSKNILTLIE LPSVNTMEEE YPRIESESTV HIGVGEPDFI LGQLYHLSSV QAPEVAKSWA ALASWAYRW GRKVVDNAS(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)EGVIKVWR KVVDRIFSLY KL SCSAYFT FLKLNAGQIP LDEDDPRLHL SHRVEQSTDD MIVMATLRLL RLLVKHAGEL RQYLEHGLET TPTAPWRGII PQL FSRLNH PEVYVRQSIC NLLCRVAQDS PHLILYPAIV GTISLSSESQ ASGNKFSTAI PTLLGNIQGE ELLVSECEGG SPPA SQDSN KDEPKSGLNE DQAMMQDCYS KIVDKLSSAN PTMVLQVQML VAELRRVTVL WDELWLGVLL QQHMYVLRRI QQLED EV(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)PHEK W FQDNYGDA IENALEKLK(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK) (UNK)YILRLEEIS PWLAAMTNTE IALPGEVSAR DTVTIHSVGG TITILPTKTK PKKLLFLGSD GKSYPYLFKG L EDLHLDER IMQFLSIVNT MFATINRQET PRFHARHYSV TPLGTRSGLI QWVDGATPLF GLYKRWQQRE AALQAQK(UNK) (UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK) (UNK)(UNK)(UNK)(UNK)(UNK)(UNK)(UNK) (UNK)(UNK)WPLHVMKA VLEELMEATP PNLLAKELWS SCTTPDE WW RVTQSYARST AVMSMVGYII GLGARHLDNV LIDMTTGEVV HIDYNVCFEK GKSLRVPEKV PFRMTQNIET ALGVTGVE G VFRLSCEQVL HIMRRGRETL LTLLEAFVYD PLVDWTAGGE AGFAGAVYGG GGQQAESKQS KREMEREITR SLFSSRVAE IKVNWFKNRD EMLVVLPKLD GSLDEYLSLQ EQLTDVEKLQ GKLLEEIEFL EGAEGVDHPS HTLQHRYSEH TQLQTQQRAV QEAIQVKLN EFEQWITHYQ AAFNNLEATQ LASLLQEIST QMDLGPPSYV PATAFLQNAG QAHLISQCEQ LEGEVGALLQ Q RRSVLRGC LEQLHHYATV ALQYPKAIFQ KHRIEQWKTW MEELICNTTV ERCQELYRKY EMQYAPQPPP TVCQFITATE MT LQRYAAD INSRLIRQVE RLKQEAVTVP VCEDQLKEIE RCIKVFLHEN GEEGSLSLAS VIISALCTLT RRNLMMEGAA SSA GEQLVD LTSRDGAWFL EELCSMSGNV TCLVQLLKQC HLVPQDLDIP NPMEASETVH LANGVYTSLQ ELNSNFRQII FPEA LRCLM KGEYTLESML HELDGLIEQT TDGVPLQTLV ESLQAYLRNA AMGLEEETHA HYIDVARLLH AQYGELIQPR NGSVD ETPK MSAGQMLLVA FDGMFAQVET AFSLLVEKLN KMEIPIAWRK IDIIREARST QVNFFDDDNH RQVLEEIFFL KRLQTI KEF FRLCGTFSKT LSGSSSLEDQ NTVNGPVQIV NVKTLFRNSC FSEDQMAKPI KAFTADFVRQ LLIGLPNQAL GLTLCSF IS ALGVDIIAQV EAKDFGAESK VSVDDLCKKA VEHNIQIGKF SQLVMNRATV LASSYDTAWK KHDLVRRLET SISSCKTS L QRVQLHIAMF QWQHEDLLIN RPQAMSVTPP PRSAILTSMK KKLHTLSQIE TSIATVQEKL AALESSIEQR LKWAGGANP ALAPVLQDFE ATIAERRNLV LKESQRASQV TFLCSNIIHF ESLRTRTAEA LNLDAALFEL IKRCQQMCSF ASQFNSSVSE LELRLLQRV DTGLEHPIGS SEWLLSAHKQ LTQDMSTQRA IQTEKEQQIE TVCETIQNLV DNIKTVLTGH NRQLGDVKHL L KAMAKDEE AALADGEDVP YENSVRQFLG EYKSWQDNIQ TVLFTLVQAM GQVRSQEHVE MLQEITPTLK ELKTQSQSIY NN LVSFASP LVTDATNECS SPTSSATYQP SFAAAVRSNT GQKTQPDVMS QNARKLIQKN LATSADTPPS TVPGTGKSVA CSP KKAVRD PKTGKAVQER NSYAVSVWKR VKAKLEGRDV DPNRRMSVAE QVDYVIKEAT NLDNLAQLYE GWTAWV UniProtKB: Serine/threonine-protein kinase SMG1, Serine/threonine-protein kinase SMG1, Serine/threonine-protein kinase SMG1, Serine/threonine-protein kinase SMG1, Serine/threonine-protein kinase SMG1 |
-分子 #2: Protein SMG8
| 分子 | 名称: Protein SMG8 / タイプ: protein_or_peptide / ID: 2 / コピー数: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 109.82575 KDa |
| 組換発現 | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MAGPVSLRDL LMGASAWMGS ESPGGSPTEG GGSAAGGPEP PWREDEICVV GIFGKTALRL NSEKFSLVNT VCDRQVFPLF RHQDPGDPG PGIRTEAGAV GEAGGAEDPG AAAGGSVRGS GAVAEGNRTE AGSQDYSLLQ AYYSQESKVL YLLLTSICDN S QLLRACRA ...文字列: MAGPVSLRDL LMGASAWMGS ESPGGSPTEG GGSAAGGPEP PWREDEICVV GIFGKTALRL NSEKFSLVNT VCDRQVFPLF RHQDPGDPG PGIRTEAGAV GEAGGAEDPG AAAGGSVRGS GAVAEGNRTE AGSQDYSLLQ AYYSQESKVL YLLLTSICDN S QLLRACRA LQSGEAGGGL SLPHAEAHEF WKHQEKLQCL SLLYLFSVCH ILLLVHPTCS FDITYDRVFR ALDGLRQKVL PL LKTAIKD CPVGKDWKLN CRPCPPRLLF LFQLNGALKV EPPRNQDPAH PDKPKKHSPK RRLQHALEDQ IYRIFRKSRV LTN QSINCL FTVPANQAFV YIVPGSQEED PVGMLLDQLR SHCTVKDPES LLVPAPLSGP RRYQVMRQHS RQQLSFHIDS SSSS SSGQL VDFTLREFLW QHVELVLSKK GFDDSVGRNP QPSHFELPTY QKWISAASKL YEVAIDGKEE DLGSPTGELT SKILS SIKV LEGFLDIDTK FSENRCQKAL PMAHSAYQSN LPHNYTMTVH KNQLAQALRV YSQHARGPAF HKYAMQLHED CYKFWS NGH QLCEERSLTD QHCVHKFHSL PKSGEKPEAD RNPPVLYHNS RARSTGACNC GRKQAPRDDP FDIKAANYDF YQLLEEK CC GKLDHINFPV FEPSTPDPAP AKNESSPAPP DSDADKLKEK EPQTQGESTS LSLALSLGQS TDSLGTYPAD PQAGGDNP E VHGQVEVKTE KRPNFVDRQA STVEYLPGML HSNCPKGLLP KFSSWSLVKL GPAKSYNFHT GLDQQGFIPG TNYLMPWDI VIRTRAEDEG DLDTNSWPAP NKAIPGKRSA VVMGRGRRRD DIARAFVGFE YEDSRGRRFM CSGPDKVMKV MGSGPKESAL KALNSDMPL YILSSSQGRG LKPHYAQLMR LFVVVPDAPL QIILMPQVQP GPPPCPVFYP EKQEITLPPD GLWVLRFPYA Y VTERGPCF PPKENVQLMS YKVLRGVLKA VTQ UniProtKB: Nonsense-mediated mRNA decay factor SMG8 |
-分子 #3: Protein SMG9
| 分子 | 名称: Protein SMG9 / タイプ: protein_or_peptide / ID: 3 / コピー数: 1 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 57.717473 KDa |
| 組換発現 | 生物種: Homo sapiens (ヒト) |
| 配列 | 文字列: MSESGHSQPG LYGIERRRRW KEPGSGGPQN LSGPGGRERD YIAPWERERR DASEETSTSV MQKTPIILSK PPAERSKQPP PPTAPAAPP APAPLEKPIV LMKPREEGKG PVAVTGASTP EGTAPPPPAA PAPPKGEKEG QRPTQPVYQI QNRGMGTAAP A AMDPVVGQ ...文字列: MSESGHSQPG LYGIERRRRW KEPGSGGPQN LSGPGGRERD YIAPWERERR DASEETSTSV MQKTPIILSK PPAERSKQPP PPTAPAAPP APAPLEKPIV LMKPREEGKG PVAVTGASTP EGTAPPPPAA PAPPKGEKEG QRPTQPVYQI QNRGMGTAAP A AMDPVVGQ AKLLPPERMK HSIKLVDDQM NWCDSAIEYL LDQTDVLVVG VLGLQGTGKS MVMSLLSANT PEEDQRTYVF RA QSAEMKE RGGNQTSGID FFITQERIVF LDTQPILSPS ILDHLINNDR KLPPEYNLPH TYVEMQSLQI AAFLFTVCHV VIV VQDWFT DLSLYRFLQT AEMVKPSTPS PSHESSSSSG SDEGTEYYPH LVFLQNKARR EDFCPRKLRQ MHLMIDQLMA HSHL RYKGT LSMLQCNVFP GLPPDFLDSE VNLFLVPFMD SEAESENPPR AGPGSSPLFS LLPGYRGHPS FQSLVSKLRS QVMSM ARPQ LSHTILTEKN WFHYAARIWD GVRKSSALAE YSRLLA UniProtKB: Nonsense-mediated mRNA decay factor SMG9 |
-分子 #4: INOSITOL HEXAKISPHOSPHATE
| 分子 | 名称: INOSITOL HEXAKISPHOSPHATE / タイプ: ligand / ID: 4 / コピー数: 1 / 式: IHP |
|---|---|
| 分子量 | 理論値: 660.035 Da |
| Chemical component information | 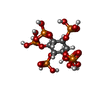 ChemComp-IHP: |
-分子 #5: ADENOSINE-5'-TRIPHOSPHATE
| 分子 | 名称: ADENOSINE-5'-TRIPHOSPHATE / タイプ: ligand / ID: 5 / コピー数: 1 / 式: ATP |
|---|---|
| 分子量 | 理論値: 507.181 Da |
| Chemical component information |  ChemComp-ATP: |
-分子 #6: MAGNESIUM ION
| 分子 | 名称: MAGNESIUM ION / タイプ: ligand / ID: 6 / コピー数: 1 / 式: MG |
|---|---|
| 分子量 | 理論値: 24.305 Da |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | particle |
-試料調製
| 緩衝液 | pH: 7.5 / 詳細: PBS |
|---|---|
| 凍結 | 凍結剤: ETHANE-PROPANE |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 検出モード: COUNTING / 平均電子線量: 52.8 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: INSILICO MODEL |
|---|---|
| 最終 再構成 | 想定した対称性 - 点群: C1 (非対称) / 解像度のタイプ: BY AUTHOR / 解像度: 3.45 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 使用した粒子像数: 214254 |
| 初期 角度割当 | タイプ: PROJECTION MATCHING |
| 最終 角度割当 | タイプ: PROJECTION MATCHING |
