 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 5fyw | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
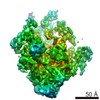
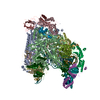
| タイトル | Transcription initiation complex structures elucidate DNA opening (OC) | |||||||||
 要素 要素 |
| |||||||||
 キーワード キーワード | TRANSCRIPTION / GENE EXPRESSION / TRANSCRIPTION INITIATION | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報RNA polymerase II complex recruiting activity / TFIIA-class transcription factor complex binding / transcription factor TFIIIB complex / RNA polymerase III preinitiation complex assembly / RNA polymerase III transcription regulatory region sequence-specific DNA binding / regulation of transcription by RNA polymerase III / RNA polymerase I general transcription initiation factor binding / transcription factor TFIIE complex / transcription open complex formation at RNA polymerase II promoter / TFIIF-class transcription factor complex binding ...RNA polymerase II complex recruiting activity / TFIIA-class transcription factor complex binding / transcription factor TFIIIB complex / RNA polymerase III preinitiation complex assembly / RNA polymerase III transcription regulatory region sequence-specific DNA binding / regulation of transcription by RNA polymerase III / RNA polymerase I general transcription initiation factor binding / transcription factor TFIIE complex / transcription open complex formation at RNA polymerase II promoter / TFIIF-class transcription factor complex binding / RPB4-RPB7 complex / transcriptional start site selection at RNA polymerase II promoter / transcription factor TFIIF complex / transcription factor TFIIA complex / RNA polymerase I preinitiation complex assembly / transcription preinitiation complex / nuclear-transcribed mRNA catabolic process, deadenylation-dependent decay / DNA binding, bending / RNA Polymerase I Transcription Initiation / Processing of Capped Intron-Containing Pre-mRNA / RNA Polymerase III Transcription Initiation From Type 2 Promoter / RNA Pol II CTD phosphorylation and interaction with CE / Formation of the Early Elongation Complex / mRNA Capping / RNA polymerase II transcribes snRNA genes / TP53 Regulates Transcription of DNA Repair Genes / termination of RNA polymerase II transcription / RNA Polymerase II Promoter Escape / RNA Polymerase II Transcription Pre-Initiation And Promoter Opening / RNA Polymerase II Transcription Initiation / RNA Polymerase II Transcription Initiation And Promoter Clearance / transcription factor TFIID complex / RNA-templated transcription / RNA Polymerase II Pre-transcription Events / RNA polymerase II general transcription initiation factor activity / termination of RNA polymerase III transcription / Formation of TC-NER Pre-Incision Complex / termination of RNA polymerase I transcription / transcription initiation at RNA polymerase III promoter / RNA Polymerase I Promoter Escape / RNA polymerase II complex binding / protein phosphatase activator activity / nucleolar large rRNA transcription by RNA polymerase I / maintenance of transcriptional fidelity during transcription elongation by RNA polymerase II / Gap-filling DNA repair synthesis and ligation in TC-NER / transcription initiation at RNA polymerase I promoter / transcription by RNA polymerase I / Estrogen-dependent gene expression / positive regulation of nuclear-transcribed mRNA poly(A) tail shortening / nuclear-transcribed mRNA catabolic process / transcription by RNA polymerase III / RNA polymerase II activity / Dual incision in TC-NER / transcription elongation by RNA polymerase I / positive regulation of transcription initiation by RNA polymerase II / tRNA transcription by RNA polymerase III / RNA polymerase II core promoter sequence-specific DNA binding / RNA polymerase I activity / RNA polymerase I complex / RNA polymerase III complex / positive regulation of translational initiation / transcription-coupled nucleotide-excision repair / RNA polymerase II, core complex / translesion synthesis / positive regulation of transcription elongation by RNA polymerase II / RNA polymerase II preinitiation complex assembly / transcription coregulator activity / translation initiation factor binding / TBP-class protein binding / DNA-templated transcription initiation / transcription initiation at RNA polymerase II promoter / transcription elongation by RNA polymerase II / P-body / ribonucleoside binding / DNA-directed 5'-3' RNA polymerase activity / DNA-directed RNA polymerase / cytoplasmic stress granule / mRNA processing / disordered domain specific binding / ribosome biogenesis / peroxisome / single-stranded DNA binding / DNA-binding transcription factor binding / transcription regulator complex / nucleic acid binding / RNA polymerase II-specific DNA-binding transcription factor binding / transcription by RNA polymerase II / single-stranded RNA binding / protein dimerization activity / negative regulation of DNA-templated transcription / chromatin binding / mRNA binding / nucleotide binding / regulation of DNA-templated transcription / nucleolus / positive regulation of transcription by RNA polymerase II / protein-containing complex / mitochondrion / DNA binding / zinc ion binding 類似検索 - 分子機能 | |||||||||
| 生物種 |  SYNTHETIC CONSTRUCT (人工物) | |||||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.35 Å | |||||||||
 データ登録者 データ登録者 | Plaschka, C. / Hantsche, M. / Dienemann, C. / Burzinski, C. / Plitzko, J. / Cramer, P. | |||||||||
 引用 引用 | ジャーナル: Nature / 年: 2016 タイトル: Transcription initiation complex structures elucidate DNA opening. 著者: C Plaschka / M Hantsche / C Dienemann / C Burzinski / J Plitzko / P Cramer /  要旨: Transcription of eukaryotic protein-coding genes begins with assembly of the RNA polymerase (Pol) II initiation complex and promoter DNA opening. Here we report cryo-electron microscopy (cryo-EM) ...Transcription of eukaryotic protein-coding genes begins with assembly of the RNA polymerase (Pol) II initiation complex and promoter DNA opening. Here we report cryo-electron microscopy (cryo-EM) structures of yeast initiation complexes containing closed and open DNA at resolutions of 8.8 Å and 3.6 Å, respectively. DNA is positioned and retained over the Pol II cleft by a network of interactions between the TATA-box-binding protein TBP and transcription factors TFIIA, TFIIB, TFIIE, and TFIIF. DNA opening occurs around the tip of the Pol II clamp and the TFIIE 'extended winged helix' domain, and can occur in the absence of TFIIH. Loading of the DNA template strand into the active centre may be facilitated by movements of obstructing protein elements triggered by allosteric binding of the TFIIE 'E-ribbon' domain. The results suggest a unified model for transcription initiation with a key event, the trapping of open promoter DNA by extended protein-protein and protein-DNA contacts. | |||||||||
| 履歴 |
| |||||||||
| Remark 700 | SHEET DETERMINATION METHOD: DSSP THE SHEETS PRESENTED AS "AM" IN EACH CHAIN ON SHEET RECORDS BELOW ... SHEET DETERMINATION METHOD: DSSP THE SHEETS PRESENTED AS "AM" IN EACH CHAIN ON SHEET RECORDS BELOW IS ACTUALLY AN 9-STRANDED BARREL THIS IS REPRESENTED BY A 10-STRANDED SHEET IN WHICH THE FIRST AND LAST STRANDS ARE IDENTICAL. THE SHEETS PRESENTED AS "GB" IN EACH CHAIN ON SHEET RECORDS BELOW IS ACTUALLY AN 5-STRANDED BARREL THIS IS REPRESENTED BY A 6-STRANDED SHEET IN WHICH THE FIRST AND LAST STRANDS ARE IDENTICAL. THE SHEETS PRESENTED AS "QC" IN EACH CHAIN ON SHEET RECORDS BELOW IS ACTUALLY AN 8-STRANDED BARREL THIS IS REPRESENTED BY A 9-STRANDED SHEET IN WHICH THE FIRST AND LAST STRANDS ARE IDENTICAL. THE SHEETS PRESENTED AS "UA" IN EACH CHAIN ON SHEET RECORDS BELOW IS ACTUALLY AN 6-STRANDED BARREL THIS IS REPRESENTED BY A 7-STRANDED SHEET IN WHICH THE FIRST AND LAST STRANDS ARE IDENTICAL. |
- 構造の表示
構造の表示
| ムービー |
ムービービューア |
|---|---|
| 構造ビューア | 分子: MolmilJmol/JSmol |
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 5fyw.cif.gz | 1.1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb5fyw.ent.gz | 852.3 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 5fyw.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/fy/5fywftp://data.pdbj.org/pub/pdb/validation_reports/fy/5fyw | HTTPS FTP |
|---|
-関連構造データ
| 関連構造データ |  3378MC  3375C  3376C  3377C  3379C  3380C  3381C  3382C  3383C  5fz5C  5ip7C  5ip9C C: 同じ文献を引用 ( M: このデータのモデリングに利用したマップデータ |
|---|---|
| 類似構造データ |


-リンク
 PDBj
PDBj












































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-DNA-DIRECTED RNA POLYMERASE II SUBUNIT ... , 7種, 7分子 ABCDGIK
| #1: タンパク質 | 分子量: 191821.578 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #2: タンパク質 | 分子量: 138937.297 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #3: タンパク質 | 分子量: 35330.457 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #4: タンパク質 | 分子量: 25451.191 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #7: タンパク質 | 分子量: 19081.053 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #9: タンパク質 | 分子量: 14308.161 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #11: タンパク質 | 分子量: 13633.493 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-DNA-DIRECTED RNA POLYMERASES I, II, AND III SUBUNIT RPABC ... , 5種, 5分子 EFHJL
| #5: タンパク質 | 分子量: 25117.094 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
|---|---|
| #6: タンパク質 | 分子量: 17931.834 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #8: タンパク質 | 分子量: 16525.363 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #10: タンパク質 | 分子量: 8290.732 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #12: タンパク質 | 分子量: 7729.969 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-タンパク質 , 2種, 2分子 MO
| #13: タンパク質 | 分子量: 38257.340 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主:  |
|---|---|
| #15: タンパク質 | 分子量: 27042.275 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
-DNA鎖 , 2種, 2分子 NT
| #14: DNA鎖 | 分子量: 22424.338 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) SYNTHETIC CONSTRUCT (人工物) |
|---|---|
| #18: DNA鎖 | 分子量: 22143.199 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) SYNTHETIC CONSTRUCT (人工物) |
-TRANSCRIPTION INITIATION FACTOR IIF SUBUNIT ... , 2種, 2分子 QR
| #16: タンパク質 | 分子量: 82320.570 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
|---|---|
| #17: タンパク質 | 分子量: 46684.492 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
-TRANSCRIPTION INITIATION FACTOR IIA ... , 2種, 2分子 UV
| #19: タンパク質 | 分子量: 32230.805 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
|---|---|
| #20: タンパク質 | 分子量: 13473.070 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
-TRANSCRIPTION INITIATION FACTOR IIE SUBUNIT ... , 2種, 2分子 WX
| #21: タンパク質 | 分子量: 54804.809 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
|---|---|
| #22: タンパク質 | 分子量: 37050.434 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
-非ポリマー , 2種, 11分子 

| #23: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 10 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 10 / 由来タイプ: 合成 / 式: Zn#24: 化合物 | ChemComp-MG / |  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: YEAST POL II TRANSCRIPTION INITIATION COMPLEX (OPEN DNA) タイプ: COMPLEX |
|---|---|
| 緩衝液 | 名称: 25 MM HEPES-KOH PH 7.5, 150 MM POTASSIUM ACETATE, 2 MM MGCL2, 5 MM DTT pH: 7.5 詳細: 25 MM HEPES-KOH PH 7.5, 150 MM POTASSIUM ACETATE, 2 MM MGCL2, 5 MM DTT |
| 試料 | 濃度: 0.3 mg/ml / 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 試料支持 | 詳細: HOLEY CARBON |
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE 詳細: CRYOGEN - ETHANE. INSTRUMENT - FEI VITROBOT MARK IV. QUANTIFOIL R 3.5- 1 HOLEY CARBON GRIDS WERE GLOW-DISCHARGED BEFORE DEPOSITION OF 4.5 MICROLITERS OF SAMPLE. GRIDS WERE THEN BLOTTED FOR 8. ...詳細: CRYOGEN - ETHANE. INSTRUMENT - FEI VITROBOT MARK IV. QUANTIFOIL R 3.5- 1 HOLEY CARBON GRIDS WERE GLOW-DISCHARGED BEFORE DEPOSITION OF 4.5 MICROLITERS OF SAMPLE. GRIDS WERE THEN BLOTTED FOR 8.5 S AND PLUNGE- FROZEN IN LIQUID ETHANE. |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS / 日付: 2016年4月26日 |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 37000 X / 倍率(補正後): 37037 X / 最大 デフォーカス(公称値): 4200 nm / 最小 デフォーカス(公称値): 700 nm / Cs: 2.3 mm |
| 撮影 | 電子線照射量: 33 e/Å2 フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
| 画像スキャン | デジタル画像の数: 1756 |
- 解析
解析
| EMソフトウェア | 名称: RELION / バージョン: 1.3 / カテゴリ: 3次元再構成 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | 詳細: EACH PARTICLE | ||||||||||||
| 対称性 | 点対称性: C1 (非対称) | ||||||||||||
| 3次元再構成 | 解像度: 4.35 Å / 粒子像の数: 11231 / ピクセルサイズ(実測値): 1.35 Å 詳細: RESIDUES INDICATED AS POLY-ALANINE ARE OF UNCERTAIN AMINO ACID REGISTER. THE FOLLOWING REGIONS WERE MODELED INTO THE INDICATED UNSHARPENED MAP DUE TO WEAKER DENSITY. RPB2 RESIDUES 919-932 ...詳細: RESIDUES INDICATED AS POLY-ALANINE ARE OF UNCERTAIN AMINO ACID REGISTER. THE FOLLOWING REGIONS WERE MODELED INTO THE INDICATED UNSHARPENED MAP DUE TO WEAKER DENSITY. RPB2 RESIDUES 919-932 (OC1, EMD-3375). TFIIF SUBUNIT TFG1 RESIDUES 327-330 (POLY-ALANINE) AND 404-415 (POLY-ALANINE) (OC4, EMD-3380). TFIIF SUBUNIT TFG2 RESIDUES 233-244 (POLY- ALANINE) (OC4, EMD-3380). TFIIE SUBUNITS TFA1 (POLY- ALANINE) AND TFA2 (POLY-ALANINE) AND TFIIF SUBUNIT TFG2 WINGED HELIX DOMAIN RESIDUES 294-352 (POLY-ALANINE) (OC3, EMD-3378 AND OC3-FOCUSED, EMD-3379). TFIIF SUBUNIT TFG2 WINGED HELIX DOMAIN RESIDUES 359-370 (UNK) WERE PUTATIVELY ASSIGNED TO TFG2 (OC3-FOCUSED, EMD-3379). TFIIE SUBUNIT TFA2 RESIDUES 256-280 SHOW UNCERTAIN CONNECTIVITY. THE SEQUENCE OF THE TFIIF TFG2 LINKER (RESIDUES 249-280) WAS ASSIGNED BASED ON THE STRONG SIDECHAIN DENSITY OF RESIDUES I252, L255, F277, L278 AND THEIR FIT TO THE LOCAL CHEMICAL ENVIRONMENT. TFIIF SUBUNIT TFG1 RESIDUES 21-35 WERE MODELED BASED ON A SEPARATE X-RAY ANALYSIS (PDB 5IP7) AND FITTED (OC4 ROUND 2 CLASS 2). SUBMISSION BASED ON EXPERIMENTAL DATA FROM EMDB EMD-3378. (DEPOSITION ID: 14360). 対称性のタイプ: POINT | ||||||||||||
| 原子モデル構築 | プロトコル: OTHER / 空間: REAL / 詳細: REFINEMENT PROTOCOL--CRYO-EM | ||||||||||||
| 精密化 | 最高解像度: 4.35 Å | ||||||||||||
| 精密化ステップ | サイクル: LAST / 最高解像度: 4.35 Å
|
