 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-4v4w: Structure of a SecM-stalled E. coli ribosome complex obtained by ... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 4v4w | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Structure of a SecM-stalled E. coli ribosome complex obtained by fitting atomic models for RNA and protein components into cryo-EM map EMD-1143 | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | RIBOSOME / SecM / nascent chain / signal transduction / RNA world / polypeptide exit tunnel / translocation / elongation arrest / protein-conducting channel | |||||||||
| Function / homology |  Function and homology information Function and homology informationstringent response / transcription antitermination factor activity, RNA binding / ornithine decarboxylase inhibitor activity / misfolded RNA binding / Group I intron splicing / RNA folding / translational termination / transcriptional attenuation / endoribonuclease inhibitor activity / positive regulation of ribosome biogenesis ...stringent response / transcription antitermination factor activity, RNA binding / ornithine decarboxylase inhibitor activity / misfolded RNA binding / Group I intron splicing / RNA folding / translational termination / transcriptional attenuation / endoribonuclease inhibitor activity / positive regulation of ribosome biogenesis / RNA-binding transcription regulator activity / four-way junction DNA binding / negative regulation of cytoplasmic translation / DnaA-L2 complex / regulation of mRNA stability / translation repressor activity / negative regulation of translational initiation / negative regulation of DNA-templated DNA replication initiation / mRNA regulatory element binding translation repressor activity / positive regulation of RNA splicing / regulation of DNA-templated transcription elongation / transcription elongation factor complex / response to reactive oxygen species / cytosolic ribosome assembly / ribosome assembly / assembly of large subunit precursor of preribosome / transcription antitermination / DNA endonuclease activity / regulation of cell growth / DNA-templated transcription termination / response to radiation / maintenance of translational fidelity / mRNA 5'-UTR binding / regulation of translation / large ribosomal subunit / transferase activity / ribosomal small subunit assembly / ribosome binding / ribosome biogenesis / ribosomal small subunit biogenesis / 5S rRNA binding / ribosomal large subunit assembly / small ribosomal subunit / small ribosomal subunit rRNA binding / large ribosomal subunit rRNA binding / cytosolic small ribosomal subunit / cytosolic large ribosomal subunit / cytoplasmic translation / tRNA binding / negative regulation of translation / rRNA binding / structural constituent of ribosome / ribosome / translation / response to antibiotic / negative regulation of DNA-templated transcription / hydrolase activity / mRNA binding / protein homodimerization activity / DNA binding / RNA binding / zinc ion binding / membrane / cytosol / cytoplasm Similarity search - Function | |||||||||
| Biological species |  | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 15 Å | |||||||||
 Authors Authors | Mitra, K. / Frank, J. | |||||||||
 Citation Citation | Journal: Mol Cell / Year: 2006 Title: Elongation arrest by SecM via a cascade of ribosomal RNA rearrangements. Authors: Kakoli Mitra / Christiane Schaffitzel / Felcy Fabiola / Michael S Chapman / Nenad Ban / Joachim Frank /  Abstract: In E. coli, the SecM nascent polypeptide causes elongation arrest, while interacting with 23S RNA bases A2058 and A749-753 in the exit tunnel of the large ribosomal subunit. We compared atomic models ...In E. coli, the SecM nascent polypeptide causes elongation arrest, while interacting with 23S RNA bases A2058 and A749-753 in the exit tunnel of the large ribosomal subunit. We compared atomic models fitted by real-space refinement into cryo-electron microscopy reconstructions of a pretranslocational and SecM-stalled E. coli ribosome complex. A cascade of RNA rearrangements propagates from the exit tunnel throughout the large subunit, affecting intersubunit bridges and tRNA positions, which in turn reorient small subunit RNA elements. Elongation arrest could result from the inhibition of mRNA.(tRNAs) translocation, E site tRNA egress, and perhaps translation factor activation at the GTPase-associated center. Our study suggests that the specific secondary and tertiary arrangement of ribosomal RNA provides the basis for internal signal transduction within the ribosome. Thus, the ribosome may itself have the ability to regulate its progression through translation by modulating its structure and consequently its receptivity to activation by cofactors. #1: Journal: Nature / Year: 2005Title: Structure of the E. coli protein-conducting channel bound to a translating ribosome Authors: Mitra, K. / Schaffitzel, C. / Shaikh, T. / Tama, F. / Jenni, S. / Brooks III, C.L. / Ban, N. / Frank, J. #2: Journal: Cell(Cambridge,Mass.) / Year: 2003Title: Locking and unlocking of ribosomal motions Authors: Valle, M. / Zavialov, A.V. / Sengupta, J. / Rawat, U. / Ehrenberg, M. / Frank, J. #3: Journal: FEBS Lett / Year: 2006 Title: A model for co-translational translocation: ribosome-regulated nascent polypeptide translocation at the protein-conducting channel. Authors: Kakoli Mitra / Joachim Frank / Abstract: The protein-conducting channel (PCC) must allow both the translocation of soluble polypeptide regions across, and the lateral partitioning of hydrophobic transmembrane helices (TMHs) into, the ...The protein-conducting channel (PCC) must allow both the translocation of soluble polypeptide regions across, and the lateral partitioning of hydrophobic transmembrane helices (TMHs) into, the membrane. We have analyzed existing structures of ribosomes and ribosome-PCC complexes and observe conformational changes suggesting that the ribosome may sense and orient the nascent polypeptide and also facilitate conformational changes in the PCC, subsequently directing the nascent polypeptide into the appropriate PCC-mediated translocation mode. The PCC is predicted to be able to accommodate one central, consolidated channel or two segregated pores with different lipid accessibilities, which may enable the lipid-mediated partitioning of a TMH from one pore, while the other, aqueous, pore allows translocation of a hydrophilic polypeptide segment. Our hypothesis suggests a plausible mechanism for the transitioning of the PCC between different configurations. | |||||||||
| History |
| |||||||||
| Remark 400 | COMPOUND THIS FILE, 2GYB, CONTAINS THE 30S SUBUNIT OF THE WHOLE RIBOSOMAL COMPLEX. THE ENTIRE CRYO- ...COMPOUND THIS FILE, 2GYB, CONTAINS THE 30S SUBUNIT OF THE WHOLE RIBOSOMAL COMPLEX. THE ENTIRE CRYO-EM STRUCTURE CONTAINS ONE 70S RIBOSOME AND ARE DEPOSITED UNDER 2GYB (30S SUBUNIT) AND 2GYC (50S SUBUNIT). | |||||||||
| Remark 999 | SEQUENCE THIS PDB FILE WAS OBTAINED UPON RIGID BODY REFINEMENT OF A STARTING PDB FILE INTO A CRYO- ...SEQUENCE THIS PDB FILE WAS OBTAINED UPON RIGID BODY REFINEMENT OF A STARTING PDB FILE INTO A CRYO-EM MAP. THE E. COLI RIBOSOMES ARE OBTAINED NATURALLY, BUT FOR THE MODELING OF THE PROTEINS INTO THE EM MAP, AUTHORS SOMETIMES DID NOT MODEL IN THE ENTIRE PROTEIN. SOME RESIDUES ARE NOT WITHIN LINK DISTANCES FOR C-N BOND. |
- Structure visualization
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
 UCSF Chimera
UCSF Chimera- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 4v4w.cif.gz | 3 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb4v4w.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 4v4w.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/v4/4v4wftp://data.pdbj.org/pub/pdb/validation_reports/v4/4v4w | HTTPS FTP |
|---|
-Related structure data
| Related structure data | 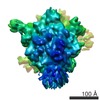 1143M  4v4vC C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |

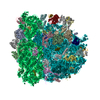
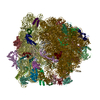
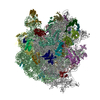
-Links
 PDBj
PDBj





























- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-RNA chain , 4 types, 6 molecules AAAUAVAWB0B9
| #1: RNA chain | Mass: 482285.656 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) | ||||
|---|---|---|---|---|---|
| #2: RNA chain | Mass: 24518.570 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Details: A-site, P-site, E-site / Source: (natural) #22: RNA chain | | Mass: 888503.500 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #23: RNA chain | | Mass: 34916.805 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
-30S ribosomal subunit protein ... , 19 types, 19 molecules ABACADAEAFAGAHAIAJAKALAMANAOAPAQARASAT
| #3: Protein | Mass: 26292.082 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|---|
| #4: Protein | Mass: 23078.785 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #5: Protein | Mass: 23311.924 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #6: Protein | Mass: 15618.051 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #7: Protein | Mass: 11181.867 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #8: Protein | Mass: 15384.665 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #9: Protein | Mass: 13857.205 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #10: Protein | Mass: 14440.777 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #11: Protein | Mass: 10996.753 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #12: Protein | Mass: 12388.068 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #13: Protein | Mass: 11245.101 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #14: Protein | Mass: 12738.911 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #15: Protein | Mass: 7117.352 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #16: Protein | Mass: 9915.350 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #17: Protein | Mass: 8822.135 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #18: Protein | Mass: 9134.767 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #19: Protein | Mass: 8178.463 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #20: Protein | Mass: 9865.542 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
| #21: Protein | Mass: 9278.929 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
+50S ribosomal protein ... , 26 types, 27 molecules B2B3B5BABBBCBDBEBFBGBHBIBJBKBLBMBNBOBQBRBSBTBUBWBXBZB1
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component |
|
|---|