 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 |  | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| タイトル | GMPCPP-Alpha1A/Beta2A-microtubule decorated with kinesin | |||||||||
 マップデータ マップデータ | ||||||||||
 試料 試料 |
| |||||||||
 キーワード キーワード | microtubule / tubulin isotype / cryo-EM structure / STRUCTURAL PROTEIN | |||||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Cilium Assembly / Sealing of the nuclear envelope (NE) by ESCRT-III / Intraflagellar transport / Carboxyterminal post-translational modifications of tubulin / cytoplasm organization / cytolytic granule membrane / COPI-independent Golgi-to-ER retrograde traffic / plus-end-directed vesicle transport along microtubule / mitocytosis ...Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Cilium Assembly / Sealing of the nuclear envelope (NE) by ESCRT-III / Intraflagellar transport / Carboxyterminal post-translational modifications of tubulin / cytoplasm organization / cytolytic granule membrane / COPI-independent Golgi-to-ER retrograde traffic / plus-end-directed vesicle transport along microtubule / mitocytosis / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / COPI-mediated anterograde transport / anterograde dendritic transport of neurotransmitter receptor complex / PKR-mediated signaling / Aggrephagy / Kinesins / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / Resolution of Sister Chromatid Cohesion / RHO GTPases activate IQGAPs / anterograde neuronal dense core vesicle transport / anterograde axonal protein transport / The role of GTSE1 in G2/M progression after G2 checkpoint / retrograde neuronal dense core vesicle transport / Recycling pathway of L1 / axonemal microtubule / COPI-dependent Golgi-to-ER retrograde traffic / RHO GTPases Activate Formins / Separation of Sister Chromatids / Hedgehog 'off' state / organelle transport along microtubule / Loss of Nlp from mitotic centrosomes / Recruitment of mitotic centrosome proteins and complexes / Loss of proteins required for interphase microtubule organization from the centrosome / Recruitment of NuMA to mitotic centrosomes / Anchoring of the basal body to the plasma membrane / glial cell differentiation / AURKA Activation by TPX2 / vesicle transport along microtubule / forebrain morphogenesis / neuron projection arborization / Regulation of PLK1 Activity at G2/M Transition / lysosome localization / cerebellar cortex morphogenesis / positive regulation of potassium ion transport / dentate gyrus development / MHC class II antigen presentation / pyramidal neuron differentiation / Kinesins / plus-end-directed microtubule motor activity / RHO GTPases activate KTN1 / stress granule disassembly / centrosome cycle / motor behavior / mitochondrion transport along microtubule / centrosome localization / COPI-dependent Golgi-to-ER retrograde traffic / microtubule motor activity / ciliary rootlet / natural killer cell mediated cytotoxicity / response to L-glutamate / kinesin complex / synaptic vesicle transport / smoothened signaling pathway / condensed chromosome / adult behavior / intercellular bridge / regulation of synapse organization / Insulin processing / microtubule-based movement / startle response / cytoplasmic microtubule / locomotory exploration behavior / microtubule polymerization / centriolar satellite / response to tumor necrosis factor / neurogenesis / response to mechanical stimulus / homeostasis of number of cells within a tissue / axon cytoplasm / phagocytic vesicle / cellular response to calcium ion / MHC class II antigen presentation / dendrite cytoplasm / positive regulation of protein localization to plasma membrane / adult locomotory behavior / regulation of membrane potential / axon guidance / locomotory behavior / positive regulation of synaptic transmission, GABAergic / hippocampus development / 加水分解酵素; 酸無水物に作用; GTPに作用・細胞または細胞小器官の運動に関与 / intracellular protein transport / synapse organization / neuron migration / visual learning / neuromuscular junction / recycling endosome / neuron differentiation / structural constituent of cytoskeleton 類似検索 - 分子機能 | |||||||||
| 生物種 |   Homo sapiens (ヒト) Homo sapiens (ヒト) | |||||||||
| 手法 | 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 4.2 Å | |||||||||
 データ登録者 データ登録者 | Zheng W / Zhao QY / Diao L / Bao L / Cong Y | |||||||||
| 資金援助 |  中国, 1件 中国, 1件
| |||||||||
 引用 引用 | ジャーナル: Acta Biochim Biophys Sin (Shanghai) / 年: 2023 タイトル: Cryo-EM of α-tubulin isotype-containing microtubules revealed a contracted structure of α4A/β2A microtubules. 著者: Lei Diao / Wei Zheng / Qiaoyu Zhao / Mingyi Liu / Zhenglin Fu / Xu Zhang / Lan Bao / Yao Cong / 要旨: Microtubules are hollow α/β-tubulin heterodimeric polymers that play critical roles in cells. In vertebrates, both α- and β-tubulins have multiple isotypes encoded by different genes, which are ...Microtubules are hollow α/β-tubulin heterodimeric polymers that play critical roles in cells. In vertebrates, both α- and β-tubulins have multiple isotypes encoded by different genes, which are intrinsic factors in regulating microtubule functions. However, the structures of microtubules composed of different tubulin isotypes, especially α-tubulin isotypes, remain largely unknown. Here, we purify recombinant tubulin heterodimers composed of different mouse α-tubulin isotypes, including α1A, α1C and α4A, with the β-tubulin isotype β2A. We further assemble and determine the cryo-electron microscopy (cryo-EM) structures of α1A/β2A, α1C/β2A, and α4A/β2A microtubules. Our structural analysis demonstrates that α4A/β2A microtubules exhibit longitudinal contraction between tubulin interdimers compared with α1A/β2A and α1C/β2A microtubules. Collectively, our findings reveal that α-tubulin isotype composition can tune microtubule structures, and also provide evidence for the "tubulin code" hypothesis. | |||||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 添付画像 |
|---|

- ダウンロードとリンク
ダウンロードとリンク
-EMDBアーカイブ
| マップデータ | emd_35790.map.gz | 46.2 MB | EMDBマップデータ形式 | |
|---|---|---|---|---|
| ヘッダ (付随情報) | emd-35790-v30.xmlemd-35790.xml | 17.9 KB 17.9 KB | 表示 表示 | EMDBヘッダ |
| 画像 |  emd_35790.png emd_35790.png | 68.7 KB | ||
| Filedesc metadata | emd-35790.cif.gz | 6.5 KB | ||
| その他 | emd_35790_half_map_1.map.gzemd_35790_half_map_2.map.gz | 146.5 MB 146.5 MB | ||
| アーカイブディレクトリ |  http://ftp.pdbj.org/pub/emdb/structures/EMD-35790ftp://ftp.pdbj.org/pub/emdb/structures/EMD-35790 http://ftp.pdbj.org/pub/emdb/structures/EMD-35790ftp://ftp.pdbj.org/pub/emdb/structures/EMD-35790 | HTTPS FTP |
-検証レポート
| 文書・要旨 | emd_35790_validation.pdf.gz | 843.6 KB | 表示 | EMDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | emd_35790_full_validation.pdf.gz | 843.1 KB | 表示 | |
| XML形式データ | emd_35790_validation.xml.gz | 18.7 KB | 表示 | |
| CIF形式データ | emd_35790_validation.cif.gz | 22.4 KB | 表示 | |
| アーカイブディレクトリ | https://ftp.pdbj.org/pub/emdb/validation_reports/EMD-35790ftp://ftp.pdbj.org/pub/emdb/validation_reports/EMD-35790 | HTTPS FTP |
-関連構造データ








-リンク
| EMDBのページ | EMDB (EBI/PDBe) / EMDataResource |
|---|---|
| 「今月の分子」の関連する項目 |


















-マップ
| ファイル | ダウンロード / ファイル: emd_35790.map.gz / 形式: CCP4 / 大きさ: 512 MB / タイプ: IMAGE STORED AS FLOATING POINT NUMBER (4 BYTES) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ボクセルのサイズ | X=Y=Z: 1.318 Å | ||||||||||||||||||||
| 密度 |
| ||||||||||||||||||||
| 対称性 | 空間群: 1 | ||||||||||||||||||||
| 詳細 | EMDB XML:
|
-添付データ
-ハーフマップ: #2
| ファイル | emd_35790_half_map_1.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
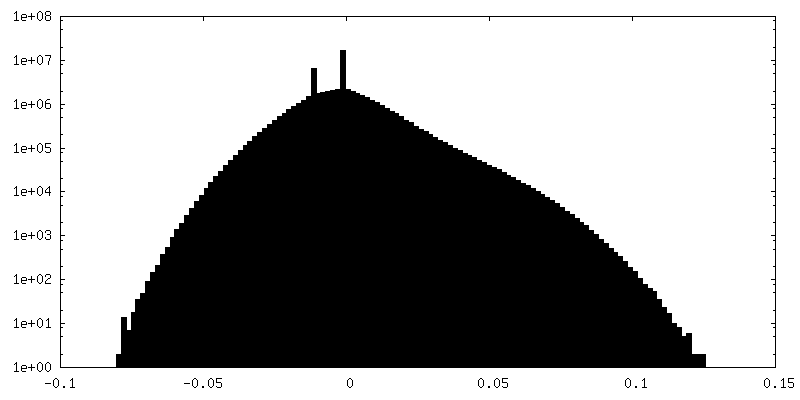
| 密度ヒストグラム |
 Z
Z Y
Y X
X






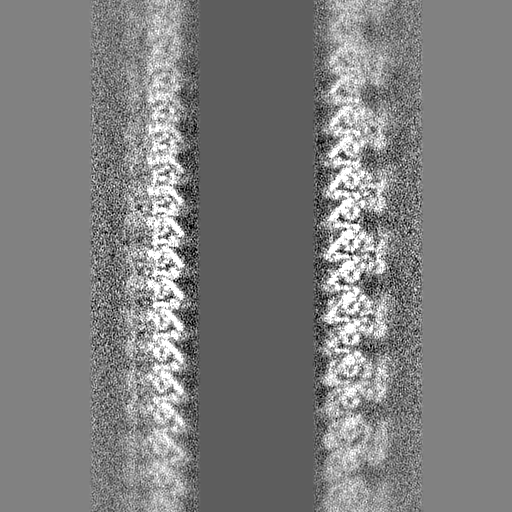
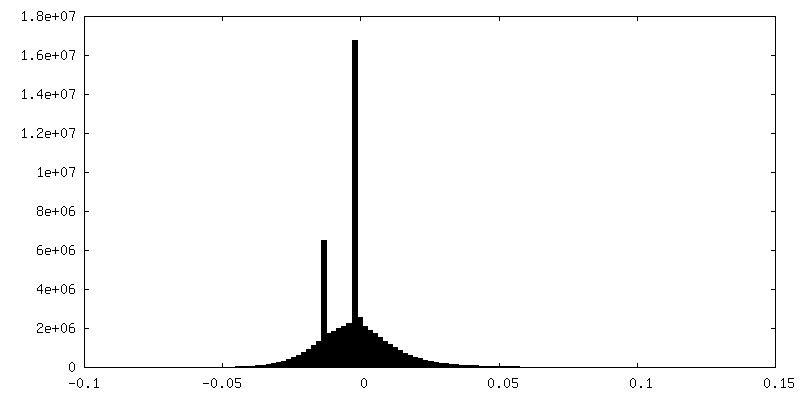
-ハーフマップ: #1
| ファイル | emd_35790_half_map_2.map | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 投影像・断面図 |
| ||||||||||||
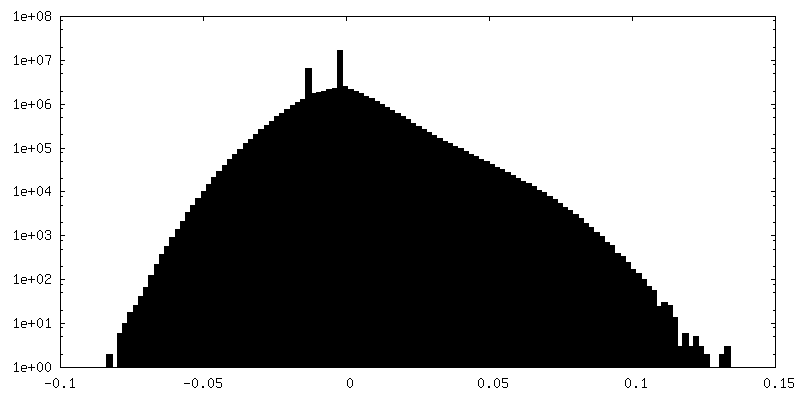
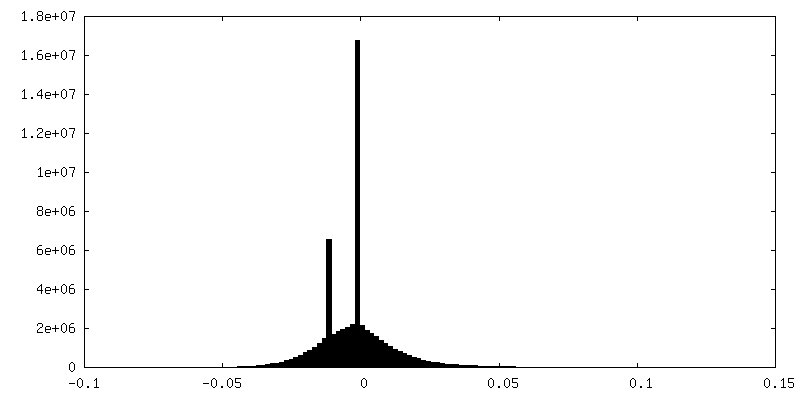
| 密度ヒストグラム |





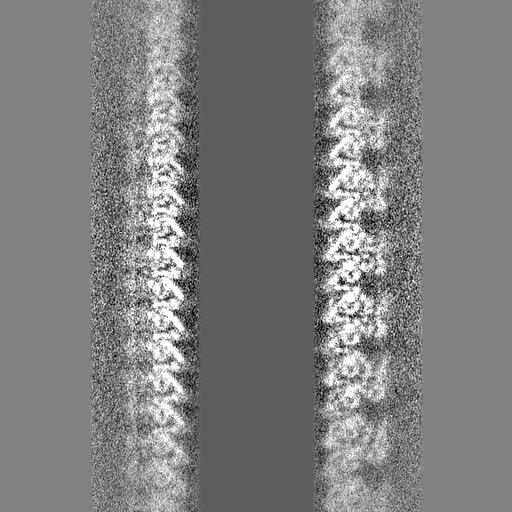
- 試料の構成要素
試料の構成要素
-全体 : GMPCPP-Alpha1A/Beta2A-microtubule decorated with kinesin
| 全体 | 名称: GMPCPP-Alpha1A/Beta2A-microtubule decorated with kinesin |
|---|---|
| 要素 |
|
-超分子 #1: GMPCPP-Alpha1A/Beta2A-microtubule decorated with kinesin
| 超分子 | 名称: GMPCPP-Alpha1A/Beta2A-microtubule decorated with kinesin タイプ: complex / ID: 1 / 親要素: 0 / 含まれる分子: #1-#3 |
|---|---|
| 由来(天然) | 生物種: |
-分子 #1: Tubulin alpha-1A chain
| 分子 | 名称: Tubulin alpha-1A chain / タイプ: protein_or_peptide / ID: 1 詳細: Author stated: the data processing is a non-standard procedure for microtubule-type of helical reconstruction with a seam, and it was performed around 2017 to 2019. We followed the procedure ...詳細: Author stated: the data processing is a non-standard procedure for microtubule-type of helical reconstruction with a seam, and it was performed around 2017 to 2019. We followed the procedure developed by Dr. Rui Zhang (R. Zhang, Cell, 2015, doi: 10.1016/j.cell.2015.07.012), with all the scripts provided by him. In this way, the generated half-maps have an applied circular mask but without touching the structural outer boundaries. コピー数: 9 / 光学異性体: LEVO EC番号: 加水分解酵素; 酸無水物に作用; GTPに作用・細胞または細胞小器官の運動に関与 |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 51.01732 KDa |
| 組換発現 | 生物種:  Insecta environmental sample (昆虫) Insecta environmental sample (昆虫) |
| 配列 | 文字列: MRECISIHVG QAGVQIGNAC WELYCLEHGI QPDGQMPSDK TIHHHHHHGG GDDSFNTFFS ETGAGKHVPR AVFVDLEPTV IDEVRTGTY RQLFHPEQLI TGKEDAANNY ARGHYTIGKE IIDLVLDRIR KLADQCTGLQ GFLVFHSFGG GTGSGFTSLL M ERLSVDYG ...文字列: MRECISIHVG QAGVQIGNAC WELYCLEHGI QPDGQMPSDK TIHHHHHHGG GDDSFNTFFS ETGAGKHVPR AVFVDLEPTV IDEVRTGTY RQLFHPEQLI TGKEDAANNY ARGHYTIGKE IIDLVLDRIR KLADQCTGLQ GFLVFHSFGG GTGSGFTSLL M ERLSVDYG KKSKLEFSIY PAPQVSTAVV EPYNSILTTH TTLEHSDCAF MVDNEAIYDI CRRNLDIERP TYTNLNRLIG QI VSSITAS LRFDGALNVD LTEFQTNLVP YPRIHFPLAT YAPVISAEKA YHEQLSVAEI TNACFEPANQ MVKCDPRHGK YMA CCLLYR GDVVPKDVNA AIATIKTKRT IQFVDWCPTG FKVGINYQPP TVVPGGDLAK VQRAVCMLSN TTAIAEAWAR LDHK FDLMY AKRAFVHWYV GEGMEEGEFS EAREDMAALE KDYEEVGVDS VEGEGEEEGE EY UniProtKB: Tubulin alpha-1A chain |
-分子 #2: Tubulin beta-2A chain
| 分子 | 名称: Tubulin beta-2A chain / タイプ: protein_or_peptide / ID: 2 / コピー数: 9 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: |
| 分子量 | 理論値: 51.150961 KDa |
| 組換発現 | 生物種: Insecta environmental sample (昆虫) |
| 配列 | 文字列: MREIVHIQAG QCGNQIGAKF WEVISDEHGI DPTGSYHGDS DLQLERINVY YNEAAGNKYV PRAILVDLEP GTMDSVRSGP FGQIFRPDN FVFGQSGAGN NWAKGHYTEG AELVDSVLDV VRKESESCDC LQGFQLTHSL GGGTGSGMGT LLISKIREEY P DRIMNTFS ...文字列: MREIVHIQAG QCGNQIGAKF WEVISDEHGI DPTGSYHGDS DLQLERINVY YNEAAGNKYV PRAILVDLEP GTMDSVRSGP FGQIFRPDN FVFGQSGAGN NWAKGHYTEG AELVDSVLDV VRKESESCDC LQGFQLTHSL GGGTGSGMGT LLISKIREEY P DRIMNTFS VMPSPKVSDT VVEPYNATLS VHQLVENTDE TYSIDNEALY DICFRTLKLT TPTYGDLNHL VSATMSGVTT CL RFPGQLN ADLRKLAVNM VPFPRLHFFM PGFAPLTSRG SQQYRALTVP ELTQQMFDSK NMMAACDPRH GRYLTVAAIF RGR MSMKEV DEQMLNVQNK NSSYFVEWIP NNVKTAVCDI PPRGLKMSAT FIGNSTAIQE LFKRISEQFT AMFRRKAFLH WYTG EGMDE MEFTEAESNM NDLVSEYQQY QDATADEQGE FEEEEGEDEA GGSGGDYKDD DK UniProtKB: Tubulin beta-2A chain |
-分子 #3: Kinesin-1 heavy chain
| 分子 | 名称: Kinesin-1 heavy chain / タイプ: protein_or_peptide / ID: 3 / コピー数: 9 / 光学異性体: LEVO |
|---|---|
| 由来(天然) | 生物種: Homo sapiens (ヒト) |
| 分子量 | 理論値: 41.721066 KDa |
| 組換発現 | 生物種:  |
| 配列 | 文字列: MGSSHHHHHH SSGLVPRGSH MASMADLAEC NIKVMCRFRP LNESEVNRGD KYIAKFQGED TVVIASKPYA FDRVFQSSTS QEQVYNDCA KKIVKDVLEG YNGTIFAYGQ TSSGKTHTME GKLHDPEGMG IIPRIVQDIF NYIYSMDENL EFHIKVSYFE I YLDKIRDL ...文字列: MGSSHHHHHH SSGLVPRGSH MASMADLAEC NIKVMCRFRP LNESEVNRGD KYIAKFQGED TVVIASKPYA FDRVFQSSTS QEQVYNDCA KKIVKDVLEG YNGTIFAYGQ TSSGKTHTME GKLHDPEGMG IIPRIVQDIF NYIYSMDENL EFHIKVSYFE I YLDKIRDL LDVSKTNLSV HEDKNRVPYV KGCTERFVCS PDEVMDTIDE GKSNRHVAVT NMNEHSSRSH SIFLINVKQE NT QTEQKLS GKLYLVDLAG SAKVSKTGAE GAVLDEAKNI NKSLSALGNV ISALAEGSTY VPYRDSKMTR ILQDSLGGNC RTT IVICCS PSSYNESETK STLLFGQRAK TIKNTVCVNV ELTAEQWKKK YEKEKE UniProtKB: Kinesin-1 heavy chain |
-分子 #4: GUANOSINE-5'-TRIPHOSPHATE
| 分子 | 名称: GUANOSINE-5'-TRIPHOSPHATE / タイプ: ligand / ID: 4 / コピー数: 9 / 式: GTP |
|---|---|
| 分子量 | 理論値: 523.18 Da |
| Chemical component information |  ChemComp-GTP: |
-分子 #5: PHOSPHOMETHYLPHOSPHONIC ACID GUANYLATE ESTER
| 分子 | 名称: PHOSPHOMETHYLPHOSPHONIC ACID GUANYLATE ESTER / タイプ: ligand / ID: 5 / コピー数: 9 / 式: G2P |
|---|---|
| 分子量 | 理論値: 521.208 Da |
| Chemical component information |  ChemComp-G2P: |
-分子 #6: ADENOSINE-5'-TRIPHOSPHATE
| 分子 | 名称: ADENOSINE-5'-TRIPHOSPHATE / タイプ: ligand / ID: 6 / コピー数: 9 / 式: ATP |
|---|---|
| 分子量 | 理論値: 507.181 Da |
| Chemical component information |  ChemComp-ATP: |
-実験情報
-構造解析
| 手法 | クライオ電子顕微鏡法 |
|---|---|
 解析 解析 | 単粒子再構成法 |
| 試料の集合状態 | filament |
-試料調製
| 緩衝液 | pH: 6.9 |
|---|---|
| 凍結 | 凍結剤: ETHANE |
- 電子顕微鏡法
電子顕微鏡法
| 顕微鏡 | FEI TITAN KRIOS |
|---|---|
| 撮影 | フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) 検出モード: COUNTING / 平均電子線量: 36.0 e/Å2 |
| 電子線 | 加速電圧: 300 kV / 電子線源:  FIELD EMISSION GUN FIELD EMISSION GUN |
| 電子光学系 | 照射モード: FLOOD BEAM / 撮影モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 1.5 µm / 最小 デフォーカス(公称値): 0.8 µm |
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
-画像解析
| 初期モデル | モデルのタイプ: EMDB MAP EMDB ID: |
|---|---|
| 最終 再構成 | 解像度のタイプ: BY AUTHOR / 解像度: 4.2 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 使用した粒子像数: 19287 |
| 初期 角度割当 | タイプ: MAXIMUM LIKELIHOOD |
| 最終 角度割当 | タイプ: MAXIMUM LIKELIHOOD |

