negative regulation of diacylglycerol biosynthetic process / protein localization to septin ring / mitotic morphogenesis checkpoint signaling / cellular bud neck septin ring / positive regulation of anaphase-promoting complex-dependent catabolic process / protein K27-linked ubiquitination / regulation of meiotic nuclear division / protein K29-linked ubiquitination / free ubiquitin chain polymerization / positive regulation of synapse maturation ...negative regulation of diacylglycerol biosynthetic process / protein localization to septin ring / mitotic morphogenesis checkpoint signaling / cellular bud neck septin ring / positive regulation of anaphase-promoting complex-dependent catabolic process / protein K27-linked ubiquitination / regulation of meiotic nuclear division / protein K29-linked ubiquitination / free ubiquitin chain polymerization / positive regulation of synapse maturation / Conversion from APC/C:Cdc20 to APC/C:Cdh1 in late anaphase / regulation of mitotic cell cycle spindle assembly checkpoint / Inactivation of APC/C via direct inhibition of the APC/C complex / APC/C:Cdc20 mediated degradation of mitotic proteins / positive regulation of synaptic plasticity / Phosphorylation of Emi1 / anaphase-promoting complex / Aberrant regulation of mitotic exit in cancer due to RB1 defects / protein branched polyubiquitination / metaphase/anaphase transition of mitotic cell cycle / regulation of meiotic cell cycle / anaphase-promoting complex-dependent catabolic process / lens fiber cell differentiation / Phosphorylation of the APC/C / anaphase-promoting complex binding / regulation of exit from mitosis / cellular bud neck / protein K6-linked ubiquitination / positive regulation of mitotic metaphase/anaphase transition / positive regulation of dendrite morphogenesis / positive regulation of ubiquitin protein ligase activity / exit from mitosis / ubiquitin ligase activator activity / protein K11-linked ubiquitination / symbiont entry into host cell via disruption of host cell glycocalyx / regulation of mitotic metaphase/anaphase transition / ubiquitin-ubiquitin ligase activity / E2 ubiquitin-conjugating enzyme / mitotic metaphase chromosome alignment / symbiont entry into host cell via disruption of host cell envelope / virus tail / mitotic G2 DNA damage checkpoint signaling / ubiquitin conjugating enzyme activity / Regulation of APC/C activators between G1/S and early anaphase / cullin family protein binding / Transcriptional Regulation by VENTX / negative regulation of cellular senescence / protein K63-linked ubiquitination / enzyme-substrate adaptor activity / APC/C:Cdc20 mediated degradation of Cyclin B / ubiquitin-like ligase-substrate adaptor activity / positive regulation of axon extension / APC-Cdc20 mediated degradation of Nek2A / protein K48-linked ubiquitination / heterochromatin / intercellular bridge / Cyclin A:Cdk2-associated events at S phase entry / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / nuclear periphery / SCF-beta-TrCP mediated degradation of Emi1 / regulation of mitotic cell cycle / Assembly of the pre-replicative complex / Cdc20:Phospho-APC/C mediated degradation of Cyclin A / APC/C:Cdh1 mediated degradation of Cdc20 and other APC/C:Cdh1 targeted proteins in late mitosis/early G1 / protein catabolic process / CDK-mediated phosphorylation and removal of Cdc6 / G protein-coupled receptor binding / brain development / kinetochore / protein modification process / spindle / G2/M transition of mitotic cell cycle / Separation of Sister Chromatids / protein polyubiquitination / neuron projection development / ubiquitin-protein transferase activity / mitotic spindle / Antigen processing: Ubiquitination & Proteasome degradation / ubiquitin protein ligase activity / nervous system development / mitotic cell cycle / Senescence-Associated Secretory Phenotype (SASP) / microtubule cytoskeleton / ubiquitin-dependent protein catabolic process / protein phosphatase binding / nuclear membrane / molecular adaptor activity / proteasome-mediated ubiquitin-dependent protein catabolic process / cell differentiation / non-specific serine/threonine protein kinase / protein ubiquitination / protein kinase activity / regulation of cell cycle / negative regulation of gene expression / cell division / protein serine kinase activity / DNA repair / intracellular membrane-bounded organelle 類似検索 - 分子機能
ジャーナル: Cell / 年: 2016 タイトル: Dual RING E3 Architectures Regulate Multiubiquitination and Ubiquitin Chain Elongation by APC/C. 著者: Nicholas G Brown / Ryan VanderLinden / Edmond R Watson / Florian Weissmann / Alban Ordureau / Kuen-Phon Wu / Wei Zhang / Shanshan Yu / Peter Y Mercredi / Joseph S Harrison / Iain F Davidson / ...著者: Nicholas G Brown / Ryan VanderLinden / Edmond R Watson / Florian Weissmann / Alban Ordureau / Kuen-Phon Wu / Wei Zhang / Shanshan Yu / Peter Y Mercredi / Joseph S Harrison / Iain F Davidson / Renping Qiao / Ying Lu / Prakash Dube / Michael R Brunner / Christy R R Grace / Darcie J Miller / David Haselbach / Marc A Jarvis / Masaya Yamaguchi / David Yanishevski / Georg Petzold / Sachdev S Sidhu / Brian Kuhlman / Marc W Kirschner / J Wade Harper / Jan-Michael Peters / Holger Stark / Brenda A Schulman / 要旨: Protein ubiquitination involves E1, E2, and E3 trienzyme cascades. E2 and RING E3 enzymes often collaborate to first prime a substrate with a single ubiquitin (UB) and then achieve different forms of ...Protein ubiquitination involves E1, E2, and E3 trienzyme cascades. E2 and RING E3 enzymes often collaborate to first prime a substrate with a single ubiquitin (UB) and then achieve different forms of polyubiquitination: multiubiquitination of several sites and elongation of linkage-specific UB chains. Here, cryo-EM and biochemistry show that the human E3 anaphase-promoting complex/cyclosome (APC/C) and its two partner E2s, UBE2C (aka UBCH10) and UBE2S, adopt specialized catalytic architectures for these two distinct forms of polyubiquitination. The APC/C RING constrains UBE2C proximal to a substrate and simultaneously binds a substrate-linked UB to drive processive multiubiquitination. Alternatively, during UB chain elongation, the RING does not bind UBE2S but rather lures an evolving substrate-linked UB to UBE2S positioned through a cullin interaction to generate a Lys11-linked chain. Our findings define mechanisms of APC/C regulation, and establish principles by which specialized E3-E2-substrate-UB architectures control different forms of polyubiquitination.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト)
Homo sapiens (ヒト) データ登録者
データ登録者 引用
引用



 構造の表示
構造の表示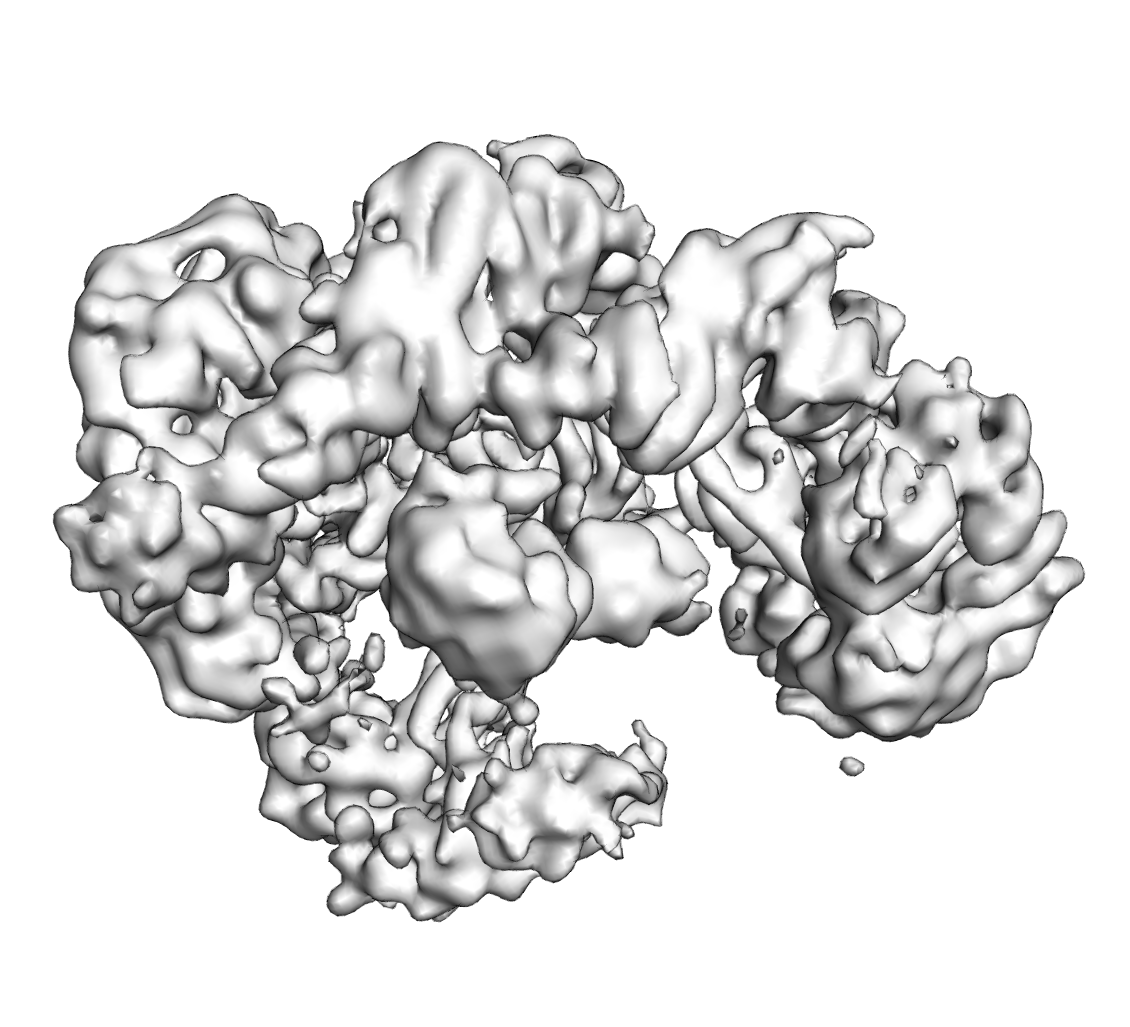
 ダウンロードとリンク
ダウンロードとリンク emd_3433.png
emd_3433.png http://ftp.pdbj.org/pub/emdb/structures/EMD-3433
http://ftp.pdbj.org/pub/emdb/structures/EMD-3433























 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





















 試料の構成要素
試料の構成要素 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN
