 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-1t1o: Components of the control 70S ribosome to provide reference for t... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 1t1o | ||||||
|---|---|---|---|---|---|---|---|
| Title | Components of the control 70S ribosome to provide reference for the RRF binding site | ||||||
 Components Components |
| ||||||
 Keywords Keywords | RIBOSOME / RRF binding position on the ribosome | ||||||
| Function / homology | RNA / RNA (> 10) Function and homology information Function and homology information | ||||||
| Biological species |  | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 12 Å | ||||||
 Authors Authors | Agrawal, R.K. / Sharma, M.R. / Kiel, M.C. / Hirokawa, G. / Booth, T.M. / Spahn, C.M. / Grassucci, R.A. / Kaji, A. / Frank, J. | ||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2004 Title: Visualization of ribosome-recycling factor on the Escherichia coli 70S ribosome: functional implications. Authors: Rajendra K Agrawal / Manjuli R Sharma / Michael C Kiel / Go Hirokawa / Timothy M Booth / Christian M T Spahn / Robert A Grassucci / Akira Kaji / Joachim Frank /  Abstract: After the termination step of protein synthesis, a deacylated tRNA and mRNA remain associated with the ribosome. The ribosome-recycling factor (RRF), together with elongation factor G (EF-G), ...After the termination step of protein synthesis, a deacylated tRNA and mRNA remain associated with the ribosome. The ribosome-recycling factor (RRF), together with elongation factor G (EF-G), disassembles this posttermination complex into mRNA, tRNA, and the ribosome. We have obtained a three-dimensional cryo-electron microscopic map of a complex of the Escherichia coli 70S ribosome and RRF. We find that RRF interacts mainly with the segments of the large ribosomal subunit's (50S) rRNA helices that are involved in the formation of two central intersubunit bridges, B2a and B3. The binding of RRF induces considerable conformational changes in some of the functional domains of the ribosome. As compared to its binding position derived previously by hydroxyl radical probing study, we find that RRF binds further inside the intersubunit space of the ribosome such that the tip of its domain I is shifted (by approximately 13 A) toward protein L5 within the central protuberance of the 50S subunit, and domain II is oriented more toward the small ribosomal subunit (30S). Overlapping binding sites of RRF, EF-G, and the P-site tRNA suggest that the binding of EF-G would trigger the removal of deacylated tRNA from the P site by moving RRF toward the ribosomal E site, and subsequent removal of mRNA may be induced by a shift in the position of 16S rRNA helix 44, which harbors part of the mRNA. #1: Journal: Cell / Year: 2001Title: High resolution structure of the large ribosomal subunit from a mesophilic eubacterium. Authors: J Harms / F Schluenzen / R Zarivach / A Bashan / S Gat / I Agmon / H Bartels / F Franceschi / A Yonath /  Abstract: We describe the high resolution structure of the large ribosomal subunit from Deinococcus radiodurans (D50S), a gram-positive mesophile suitable for binding of antibiotics and functionally relevant ...We describe the high resolution structure of the large ribosomal subunit from Deinococcus radiodurans (D50S), a gram-positive mesophile suitable for binding of antibiotics and functionally relevant ligands. The over-all structure of D50S is similar to that from the archae bacterium Haloarcula marismortui (H50S); however, a detailed comparison revealed significant differences, for example, in the orientation of nucleotides in peptidyl transferase center and in the structures of many ribosomal proteins. Analysis of ribosomal features involved in dynamic aspects of protein biosynthesis that are partially or fully disordered in H50S revealed the conformations of intersubunit bridges in unbound subunits, suggesting how they may change upon subunit association and how movements of the L1-stalk may facilitate the exit of tRNA. #2: Journal: Nature / Year: 2000Title: Structure of the 30S ribosomal subunit. Authors: B T Wimberly / D E Brodersen / W M Clemons / R J Morgan-Warren / A P Carter / C Vonrhein / T Hartsch / V Ramakrishnan /  Abstract: Genetic information encoded in messenger RNA is translated into protein by the ribosome, which is a large nucleoprotein complex comprising two subunits, denoted 30S and 50S in bacteria. Here we ...Genetic information encoded in messenger RNA is translated into protein by the ribosome, which is a large nucleoprotein complex comprising two subunits, denoted 30S and 50S in bacteria. Here we report the crystal structure of the 30S subunit from Thermus thermophilus, refined to 3 A resolution. The final atomic model rationalizes over four decades of biochemical data on the ribosome, and provides a wealth of information about RNA and protein structure, protein-RNA interactions and ribosome assembly. It is also a structural basis for analysis of the functions of the 30S subunit, such as decoding, and for understanding the action of antibiotics. The structure will facilitate the interpretation in molecular terms of lower resolution structural data on several functional states of the ribosome from electron microscopy and crystallography. | ||||||
| History |
|
- Structure visualization
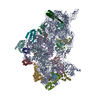
Structure visualization
| Movie |
Movie viewer |
|---|---|
| Structure viewer | Molecule: MolmilJmol/JSmol |
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 1t1o.cif.gz | 9.9 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb1t1o.ent.gz | 4.7 KB | Display | PDB format |
| PDBx/mmJSON format | 1t1o.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Summary document | 1t1o_validation.pdf.gz | 713.1 KB | Display | wwPDB validaton report |
|---|---|---|---|---|
| Full document | 1t1o_full_validation.pdf.gz | 712.6 KB | Display | |
| Data in XML | 1t1o_validation.xml.gz | 7.5 KB | Display | |
| Data in CIF | 1t1o_validation.cif.gz | 9.8 KB | Display | |
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/t1/1t1oftp://data.pdbj.org/pub/pdb/validation_reports/t1/1t1o | HTTPS FTP |
-Related structure data
| Related structure data | 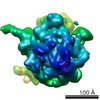 1077MC  1t1mC  1kc9 C: citing same article ( M: map data used to model this data |
|---|---|
| Similar structure data |
-Links
 PDBj
PDBj





























- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
| #1: RNA chain | Mass: 3852.344 Da / Num. of mol.: 1 / Fragment: Apical loop of Helix 43 / Source method: isolated from a natural source / Details: fitted into the cryo-EM map of the 70S ribosome / Source: (natural) |
|---|---|
| #2: RNA chain | Mass: 6077.673 Da / Num. of mol.: 1 / Fragment: Helix 69 / Source method: isolated from a natural source / Details: fitted into the cryo-EM map of the 70S ribosome / Source: (natural) |
| #3: RNA chain | Mass: 13581.124 Da / Num. of mol.: 1 / Fragment: Top portion of helix 44 / Source method: isolated from a natural source / Details: fitted into the cryo-EM map of the 70S ribosome / Source: (natural) |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: 70S-RRF complex / Type: RIBOSOME |
|---|---|
| Buffer solution | pH: 7.5 |
| Specimen | Conc.: 32 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Details: Quantifoil holley-carbon film grids |
| Vitrification | Instrument: HOMEMADE PLUNGER / Cryogen name: ETHANE / Details: Rapid-freezing in liquid ethane |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Tecnai F20 / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TECNAI F20 / Date: Jun 1, 2002 |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 200 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 50000 X / Calibrated magnification: 49696 X / Nominal defocus max: 4400 nm / Nominal defocus min: 1400 nm / Cs: 2 mm |
| Specimen holder | Temperature: 93 K / Tilt angle max: 0 ° / Tilt angle min: 0 ° |
| Image recording | Electron dose: 20 e/Å2 / Film or detector model: KODAK SO-163 FILM |
- Processing
Processing
| CTF correction | Details: CTF correction of 3D-maps by Wiener filtration | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Symmetry | Point symmetry: C1 (asymmetric) | |||||||||||||||||||||
| 3D reconstruction | Method: 3D projection matching; conjugate gradients with regularization Resolution: 12 Å / Num. of particles: 51217 / Actual pixel size: 2.82 Å / Magnification calibration: TMV / Details: SPIDER package / Symmetry type: POINT | |||||||||||||||||||||
| Atomic model building | Protocol: OTHER Details: METHOD--Cross corelation coefficient based manual fitting in O | |||||||||||||||||||||
| Atomic model building |
| |||||||||||||||||||||
| Refinement step | Cycle: LAST /
|

