[histone H3]-lysine4 N-methyltransferase / histone H3K4 monomethyltransferase activity / histone H3K4 trimethyltransferase activity / histone H3Q5ser reader activity / histone H3K4me1 reader activity / Loss of Function of KMT2D in MLL4 Complex Formation in Kabuki Syndrome / Epigenetic regulation of gene expression by MLL3 and MLL4 complexes / MLL3/4 complex / Set1C/COMPASS complex / MLL1/2 complex ...[histone H3]-lysine4 N-methyltransferase / histone H3K4 monomethyltransferase activity / histone H3K4 trimethyltransferase activity / histone H3Q5ser reader activity / histone H3K4me1 reader activity / Loss of Function of KMT2D in MLL4 Complex Formation in Kabuki Syndrome / Epigenetic regulation of gene expression by MLL3 and MLL4 complexes / MLL3/4 complex / Set1C/COMPASS complex / MLL1/2 complex / ATAC complex / NSL complex / histone H3K4 methyltransferase activity / Cardiogenesis / histone methyltransferase activity / Formation of the ternary complex, and subsequently, the 43S complex / Formation of WDR5-containing histone-modifying complexes / acyltransferase activity / Ribosomal scanning and start codon recognition / Translation initiation complex formation / histone methyltransferase complex / hemopoiesis / MLL1 complex / SARS-CoV-1 modulates host translation machinery / regulation of embryonic development / regulation of cell division / histone acetyltransferase complex / Peptide chain elongation / Selenocysteine synthesis / Formation of a pool of free 40S subunits / Eukaryotic Translation Termination / SRP-dependent cotranslational protein targeting to membrane / Response of EIF2AK4 (GCN2) to amino acid deficiency / Viral mRNA Translation / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / GTP hydrolysis and joining of the 60S ribosomal subunit / L13a-mediated translational silencing of Ceruloplasmin expression / Major pathway of rRNA processing in the nucleolus and cytosol / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / Constitutive Signaling by NOTCH1 HD Domain Mutants / IRAK2 mediated activation of TAK1 complex / Prevention of phagosomal-lysosomal fusion / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Endosomal Sorting Complex Required For Transport (ESCRT) / Membrane binding and targetting of GAG proteins / Negative regulation of FLT3 / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / positive regulation of gluconeogenesis / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / transcription initiation-coupled chromatin remodeling / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / Regulation of FZD by ubiquitination / Downregulation of ERBB4 signaling / cytosolic ribosome / APC-Cdc20 mediated degradation of Nek2A / p75NTR recruits signalling complexes / sperm principal piece / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / Regulation of pyruvate metabolism / TRAF6-mediated induction of TAK1 complex within TLR4 complex / NF-kB is activated and signals survival / Regulation of innate immune responses to cytosolic DNA / Pexophagy / Downregulation of ERBB2:ERBB3 signaling / NRIF signals cell death from the nucleus / Activated NOTCH1 Transmits Signal to the Nucleus / Regulation of PTEN localization / VLDLR internalisation and degradation / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Translesion synthesis by REV1 / Regulation of BACH1 activity / TICAM1, RIP1-mediated IKK complex recruitment / MAP3K8 (TPL2)-dependent MAPK1/3 activation / Translesion synthesis by POLK / InlB-mediated entry of Listeria monocytogenes into host cell / Degradation of CDH1 / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Josephin domain DUBs / Downregulation of TGF-beta receptor signaling / Translesion synthesis by POLI / Gap-filling DNA repair synthesis and ligation in GG-NER / Degradation of CRY and PER proteins / IKK complex recruitment mediated by RIP1 / Regulation of activated PAK-2p34 by proteasome mediated degradation / gluconeogenesis 類似検索 - 分子機能
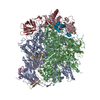
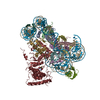
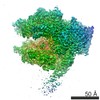
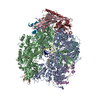
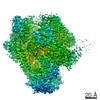
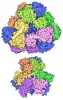
ジャーナル: Nature / 年: 2019 タイトル: Structural basis of nucleosome recognition and modification by MLL methyltransferases. 著者: Han Xue / Tonghui Yao / Mi Cao / Guanjun Zhu / Yan Li / Guiyong Yuan / Yong Chen / Ming Lei / Jing Huang / 要旨: Methyltransferases of the mixed-lineage leukaemia (MLL) family-which include MLL1, MLL2, MLL3, MLL4, SET1A and SET1B-implement methylation of histone H3 on lysine 4 (H3K4), and have critical and ...Methyltransferases of the mixed-lineage leukaemia (MLL) family-which include MLL1, MLL2, MLL3, MLL4, SET1A and SET1B-implement methylation of histone H3 on lysine 4 (H3K4), and have critical and distinct roles in the regulation of transcription in haematopoiesis, adipogenesis and development. The C-terminal catalytic SET (Su(var.)3-9, enhancer of zeste and trithorax) domains of MLL proteins are associated with a common set of regulatory factors (WDR5, RBBP5, ASH2L and DPY30) to achieve specific activities. Current knowledge of the regulation of MLL activity is limited to the catalysis of histone H3 peptides, and how H3K4 methyl marks are deposited on nucleosomes is poorly understood. H3K4 methylation is stimulated by mono-ubiquitination of histone H2B on lysine 120 (H2BK120ub1), a prevalent histone H2B mark that disrupts chromatin compaction and favours open chromatin structures, but the underlying mechanism remains unknown. Here we report cryo-electron microscopy structures of human MLL1 and MLL3 catalytic modules associated with nucleosome core particles that contain H2BK120ub1 or unmodified H2BK120. These structures demonstrate that the MLL1 and MLL3 complexes both make extensive contacts with the histone-fold and DNA regions of the nucleosome; this allows ease of access to the histone H3 tail, which is essential for the efficient methylation of H3K4. The H2B-conjugated ubiquitin binds directly to RBBP5, orienting the association between MLL1 or MLL3 and the nucleosome. The MLL1 and MLL3 complexes display different structural organizations at the interface between the WDR5, RBBP5 and MLL1 (or the corresponding MLL3) subunits, which accounts for the opposite roles of WDR5 in regulating the activity of the two enzymes. These findings transform our understanding of the structural basis for the regulation of MLL activity at the nucleosome level, and highlight the pivotal role of nucleosome regulation in histone-tail modification.
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 マップデータ
マップデータ 試料
試料 キーワード
キーワード 機能・相同性情報
機能・相同性情報 Homo sapiens (ヒト) /
Homo sapiens (ヒト) /  データ登録者
データ登録者 中国, 1件
中国, 1件  引用
引用 構造の表示
構造の表示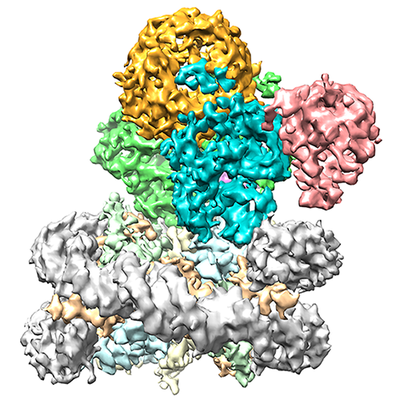
 ダウンロードとリンク
ダウンロードとリンク emd_0693.png
emd_0693.png http://ftp.pdbj.org/pub/emdb/structures/EMD-0693
http://ftp.pdbj.org/pub/emdb/structures/EMD-0693

































 Z (Sec.)
Z (Sec.) Y (Row.)
Y (Row.) X (Col.)
X (Col.)





























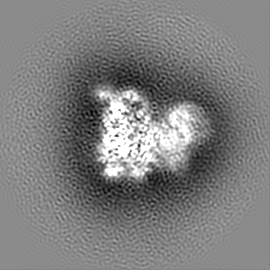
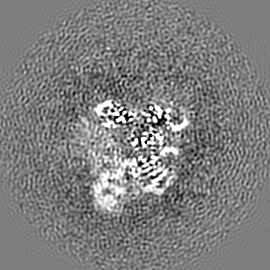
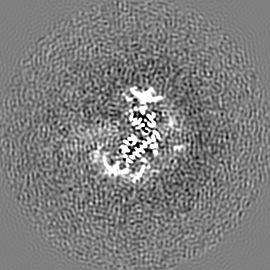
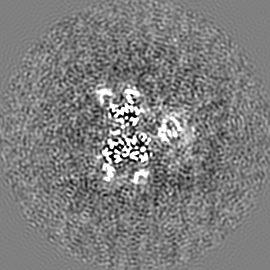

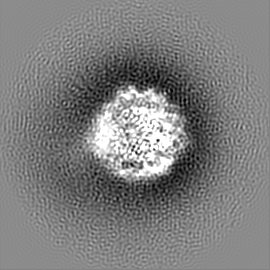
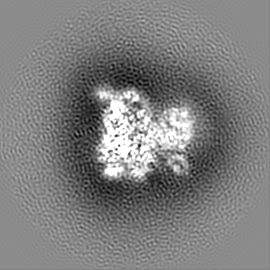


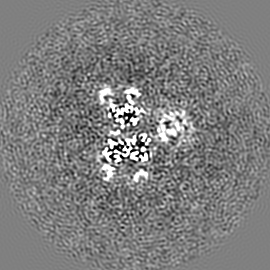

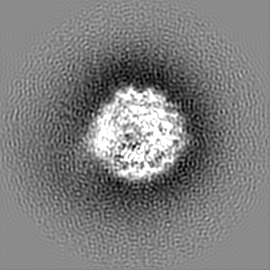

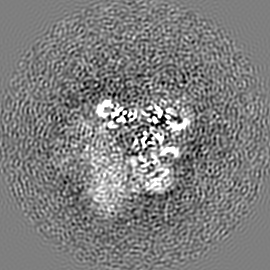
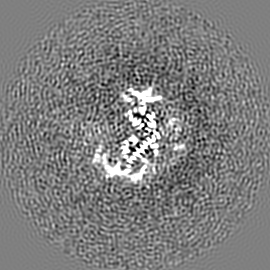
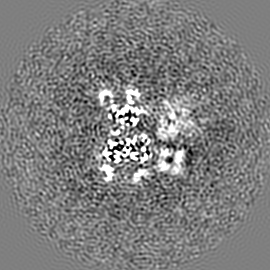

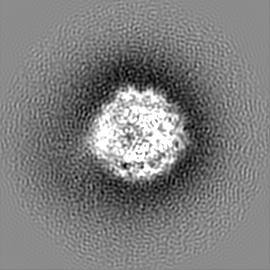

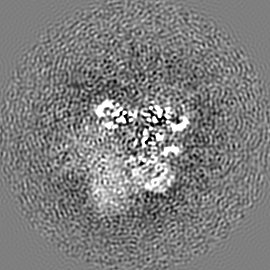
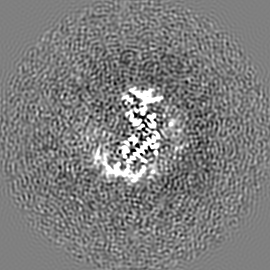
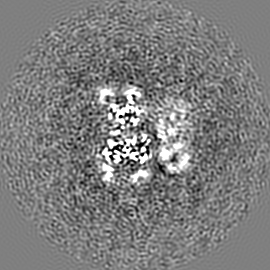

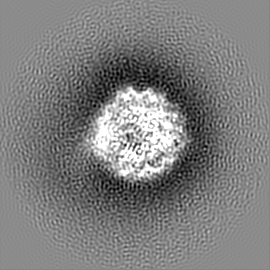
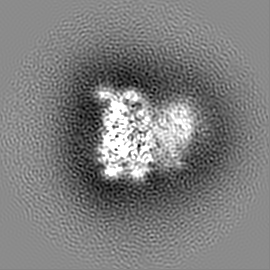




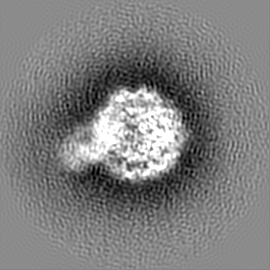

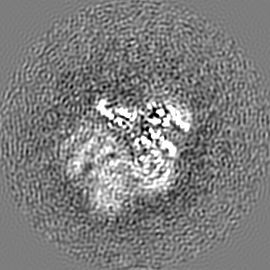
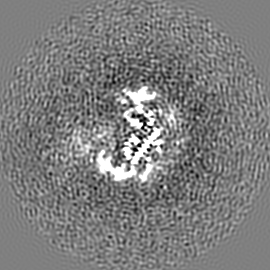

 試料の構成要素
試料の構成要素
 解析
解析 電子顕微鏡法
電子顕微鏡法 FIELD EMISSION GUN
FIELD EMISSION GUN