 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8uhg | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Structure of paused transcription complex Pol II-DSIF-NELF - poised post-translocated | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | TRANSCRIPTION/DNA/RNA /  Nucleic acids (核酸) / transcription (転写 (生物学)) / RNA polymerase II (RNAポリメラーゼII) / NELF / DSIF / pausing / TRANSCRIPTION-DNA-RNA complex Nucleic acids (核酸) / transcription (転写 (生物学)) / RNA polymerase II (RNAポリメラーゼII) / NELF / DSIF / pausing / TRANSCRIPTION-DNA-RNA complex | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報NELF complex / positive regulation of protein modification process / NTRK3 as a dependence receptor / negative regulation of DNA-templated transcription, elongation / DSIF complex / regulation of transcription elongation by RNA polymerase II / B-WICH complex positively regulates rRNA expression / RNA Polymerase I Transcription Initiation / RNA Polymerase I Promoter Escape / RNA Polymerase I Transcription Termination ...NELF complex / positive regulation of protein modification process / NTRK3 as a dependence receptor / negative regulation of DNA-templated transcription, elongation / DSIF complex / regulation of transcription elongation by RNA polymerase II / B-WICH complex positively regulates rRNA expression / RNA Polymerase I Transcription Initiation / RNA Polymerase I Promoter Escape / RNA Polymerase I Transcription Termination / RNA Polymerase III Transcription Initiation From Type 1 Promoter / RNA Polymerase III Transcription Initiation From Type 2 Promoter / RNA Polymerase III Transcription Initiation From Type 3 Promoter / Formation of RNA Pol II elongation complex / Formation of the Early Elongation Complex / Transcriptional regulation by small RNAs / RNA Polymerase II Pre-transcription Events / TP53 Regulates Transcription of DNA Repair Genes / FGFR2 alternative splicing / RNA polymerase II transcribes snRNA genes / mRNA Capping / mRNA Splicing - Major Pathway / mRNA Splicing - Minor Pathway / Processing of Capped Intron-Containing Pre-mRNA / RNA Polymerase II Promoter Escape / RNA Polymerase II Transcription Pre-Initiation And Promoter Opening / RNA Polymerase II Transcription Initiation / RNA Polymerase II Transcription Elongation / RNA Polymerase II Transcription Initiation And Promoter Clearance / RNA Pol II CTD phosphorylation and interaction with CE / Estrogen-dependent gene expression / Formation of TC-NER Pre-Incision Complex / Dual incision in TC-NER / Gap-filling DNA repair synthesis and ligation in TC-NER / negative regulation of stem cell differentiation / Abortive elongation of HIV-1 transcript in the absence of Tat / positive regulation of DNA-templated transcription, elongation / transcription elongation-coupled chromatin remodeling / RNA polymerase complex / RNA Pol II CTD phosphorylation and interaction with CE during HIV infection / RNA Pol II CTD phosphorylation and interaction with CE / Formation of the Early Elongation Complex / Formation of the HIV-1 Early Elongation Complex / mRNA Capping / : / negative regulation of transcription elongation by RNA polymerase II / maintenance of transcriptional fidelity during transcription elongation by RNA polymerase II / organelle membrane / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / positive regulation of nuclear-transcribed mRNA poly(A) tail shortening / positive regulation of macroautophagy / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / RNA polymerase II transcribes snRNA genes / tRNA transcription by RNA polymerase III / RNA polymerase III activity / RNA polymerase I activity / Tat-mediated elongation of the HIV-1 transcript / positive regulation of translational initiation / Formation of HIV-1 elongation complex containing HIV-1 Tat / RNA polymerase I complex / RNA polymerase III complex / transcription-coupled nucleotide-excision repair / localization / RNA polymerase II activity / Formation of HIV elongation complex in the absence of HIV Tat / RNA polymerase II, core complex / RNA Polymerase II Transcription Elongation / Formation of RNA Pol II elongation complex / DNA-directed RNA polymerase complex / translation initiation factor binding / RNA Polymerase II Pre-transcription Events / DNA-templated transcription initiation / transcription elongation by RNA polymerase II / 細胞分化 / transcription initiation at RNA polymerase II promoter / TP53 Regulates Transcription of DNA Repair Genes / P-body / ribonucleoside binding / 核小体 / DNA-directed 5'-3' RNA polymerase activity / ポリメラーゼ / single-stranded DNA binding / cell population proliferation / transcription by RNA polymerase II / nucleic acid binding / molecular adaptor activity / single-stranded RNA binding / positive regulation of ERK1 and ERK2 cascade / nuclear body / protein dimerization activity / protein heterodimerization activity / nucleotide binding / mRNA binding / DNA-templated transcription / chromatin binding / クロマチン / 核小体 / enzyme binding類似検索 - 分子機能 | ||||||
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) Sus scrofa (ブタ) Sus scrofa (ブタ)synthetic construct (人工物) | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.7 Å | ||||||
 データ登録者 データ登録者 | Vos, S.M. / Su, B.G. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2024 タイトル: Distinct negative elongation factor conformations regulate RNA polymerase II promoter-proximal pausing. 著者: Bonnie G Su / Seychelle M Vos / 要旨: Metazoan gene expression regulation involves pausing of RNA polymerase (Pol II) in the promoter-proximal region of genes and is stabilized by DSIF and NELF. Upon depletion of elongation factors, NELF ...Metazoan gene expression regulation involves pausing of RNA polymerase (Pol II) in the promoter-proximal region of genes and is stabilized by DSIF and NELF. Upon depletion of elongation factors, NELF appears to accompany elongating Pol II past pause sites; however, prior work indicates that NELF prevents Pol II elongation. Here, we report cryoelectron microscopy structures of Pol II-DSIF-NELF complexes with NELF in two distinct conformations corresponding to paused and poised states. The paused NELF state supports Pol II stalling, whereas the poised NELF state enables transcription elongation as it does not support a tilted RNA-DNA hybrid. Further, the poised NELF state can accommodate TFIIS binding to Pol II, allowing for Pol II reactivation at paused or backtracking sites. Finally, we observe that the NELF-A tentacle interacts with the RPB2 protrusion and is necessary for pausing. Our results define how NELF can support pausing, reactivation, and elongation by Pol II. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8uhg.cif.gz | 1.9 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8uhg.ent.gz | 1.5 MB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8uhg.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/uh/8uhgftp://data.pdbj.org/pub/pdb/validation_reports/uh/8uhg | HTTPS FTP |
|---|
-関連構造データ



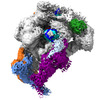

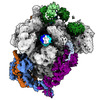


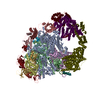

-リンク
 PDBj
PDBj



































































- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-Negative elongation factor ... , 4種, 4分子 XVWU
| #1: タンパク質 | 分子量: 43320.922 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NELFE, RD, RDBP / 発現宿主:  Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P18615 Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: P18615 |
|---|---|
| #2: タンパク質 | 分子量: 63233.250 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NELFB, COBRA1, KIAA1182 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q8WX92 |
| #3: タンパク質 | 分子量: 66315.352 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NELFCD, NELFD, TH1, TH1L, HSPC130 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q8IXH7 |
| #19: タンパク質 | 分子量: 57343.598 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: NELFA, WHSC2, P/OKcl.15 / 発現宿主: Trichoplusia ni (イラクサキンウワバ) / 参照: UniProt: Q9H3P2 |
-DNA-directed RNA polymerase ... , 7種, 7分子 ABCEGIK
| #4: タンパク質 | ポリメラーゼ 分子量: 217450.078 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A8D1DPV6, ポリメラーゼ |
|---|---|
| #5: タンパク質 | ポリメラーゼ 分子量: 134041.422 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A4X1TVZ5, ポリメラーゼ |
| #6: タンパク質 | ポリメラーゼ 分子量: 30997.557 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A481DF93 |
| #8: タンパク質 | ポリメラーゼ / RPB5 分子量: 24644.318 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A4X1VTX4 |
| #10: タンパク質 | ポリメラーゼ 分子量: 19227.205 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A4X1VKG7 |
| #12: タンパク質 | ポリメラーゼ / RNA polymerase II subunit B9 / DNA-directed RNA polymerase II subunit I / RNA polymerase II 14.5 ...RNA polymerase II subunit B9 / DNA-directed RNA polymerase II subunit I / RNA polymerase II 14.5 kDa subunit / RPB14.5 分子量: 14541.221 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: P60899 |
| #14: タンパク質 | ポリメラーゼ 分子量: 13068.013 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: F1RKE4 |
-RNA polymerase ... , 2種, 2分子 DL
| #7: タンパク質 | 分子量: 16331.255 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A8D0KES4 |
|---|---|
| #15: タンパク質 | RNAポリメラーゼII 分子量: 7018.244 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A4X1TRS6 |
-DNA-directed RNA polymerases I, II, and III subunit ... , 3種, 3分子 FHJ
| #9: タンパク質 | RNAポリメラーゼ / DNA-directed RNA polymerase II subunit F / RPB6 homolog 分子量: 14477.001 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A4X1VEK9 |
|---|---|
| #11: タンパク質 | RNAポリメラーゼ 分子量: 17162.273 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A8D1AQR7 |
| #13: タンパク質 | RNAポリメラーゼ 分子量: 7655.123 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) Sus scrofa (ブタ) / 参照: UniProt: A0A8W4F9W9 |
-DNA鎖 , 2種, 2分子 NT
| #16: DNA鎖 | 分子量: 15409.860 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|---|
| #18: DNA鎖 | 分子量: 11728.553 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
-RNA鎖 / タンパク質 , 2種, 2分子 PZ
| #17: RNA鎖 | リボ核酸 分子量: 5077.007 Da / 分子数: 1 / 由来タイプ: 合成 / 由来: (合成) synthetic construct (人工物) |
|---|---|
| #20: タンパク質 | 分子量: 121145.477 Da / 分子数: 1 / 由来タイプ: 組換発現 / 由来: (組換発現) Homo sapiens (ヒト) / 遺伝子: SUPT5H, SPT5, SPT5H / 発現宿主:  Escherichia coli (大腸菌) / 参照: UniProt: O00267 Escherichia coli (大腸菌) / 参照: UniProt: O00267 |
-非ポリマー , 2種, 9分子 

| #21: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Zn#22: 化合物 | ChemComp-MG / |  分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 1 / 由来タイプ: 合成 / 式: Mg |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: RNA Polymerase in complex with DSIF and NELFRNAポリメラーゼ タイプ: COMPLEX / Entity ID: #1-#4, #6-#20 / 由来: NATURAL |
|---|---|
| 分子量 | 実験値: NO |
| 由来(天然) | 生物種: Sus scrofa (ブタ) |
| 緩衝液 | pH: 7.4 |
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES |
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源: FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELDBright-field microscopy / 最大 デフォーカス(公称値): 2500 nm / 最小 デフォーカス(公称値): 500 nm |
| 撮影 | 電子線照射量: 51 e/Å2 フィルム・検出器のモデル: GATAN K3 BIOQUANTUM (6k x 4k) |
- 解析
解析
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 3次元再構成 | 解像度: 2.7 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 90283 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 拘束条件 |
|
