regulation of dolichol biosynthetic process / RSP5-BUL ubiquitin ligase complex / regulation of ribosomal large subunit export from nucleus / regulation of tRNA processing / regulation of tRNA export from nucleus / regulation of ubiquinone biosynthetic process / regulation of phosphate metabolic process / regulation of ergosterol biosynthetic process / protein catabolic process => GO:0030163 / fungal-type vacuole lumen ...regulation of dolichol biosynthetic process / RSP5-BUL ubiquitin ligase complex / regulation of ribosomal large subunit export from nucleus / regulation of tRNA processing / regulation of tRNA export from nucleus / regulation of ubiquinone biosynthetic process / regulation of phosphate metabolic process / regulation of ergosterol biosynthetic process / protein catabolic process => GO:0030163 / fungal-type vacuole lumen / RHOQ GTPase cycle / RHOU GTPase cycle / positive regulation of ubiquitin-dependent endocytosis / regulation of multivesicular body size / cellular response to L-arginine / ubiquitin-dependent endocytosis / ribophagy / mitochondria-associated ubiquitin-dependent protein catabolic process / free ubiquitin chain polymerization / regulation of mRNA export from nucleus / regulation of rRNA processing / endosome transport via multivesicular body sorting pathway / cellular bud tip / protein transport to vacuole involved in ubiquitin-dependent protein catabolic process via the multivesicular body sorting pathway / late endosome to vacuole transport via multivesicular body sorting pathway / actin cortical patch / positive regulation of fatty acid biosynthetic process / regulation of nitrogen utilization / ubiquitin-dependent protein catabolic process via the multivesicular body sorting pathway / HECT-type E3 ubiquitin transferase / fungal-type vacuole / nonfunctional rRNA decay / ubiquitin-ubiquitin ligase activity / poly(A)+ mRNA export from nucleus / fungal-type vacuole membrane / Regulation of PTEN stability and activity / protein quality control for misfolded or incompletely synthesized proteins / positive regulation of endocytosis / Antigen processing: Ubiquitination & Proteasome degradation / protein K63-linked ubiquitination / ubiquitin ligase complex / cytosolic ribosome / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / Prevention of phagosomal-lysosomal fusion / IRAK2 mediated activation of TAK1 complex / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Membrane binding and targetting of GAG proteins / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / Endosomal Sorting Complex Required For Transport (ESCRT) / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Negative regulation of FLT3 / Constitutive Signaling by NOTCH1 HD Domain Mutants / Regulation of FZD by ubiquitination / TICAM1,TRAF6-dependent induction of TAK1 complex / TICAM1-dependent activation of IRF3/IRF7 / NOTCH2 Activation and Transmission of Signal to the Nucleus / APC/C:Cdc20 mediated degradation of Cyclin B / p75NTR recruits signalling complexes / Downregulation of ERBB4 signaling / NOTCH3 Activation and Transmission of Signal to the Nucleus / APC-Cdc20 mediated degradation of Nek2A / TRAF6-mediated induction of TAK1 complex within TLR4 complex / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / PINK1-PRKN Mediated Mitophagy / Pexophagy / Regulation of innate immune responses to cytosolic DNA / InlA-mediated entry of Listeria monocytogenes into host cells / VLDLR internalisation and degradation / Regulation of pyruvate metabolism / Downregulation of ERBB2:ERBB3 signaling / NF-kB is activated and signals survival / Regulation of PTEN localization / NRIF signals cell death from the nucleus / Regulation of BACH1 activity / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Translesion synthesis by REV1 / phosphatidylinositol binding / MAP3K8 (TPL2)-dependent MAPK1/3 activation / Translesion synthesis by POLK / TICAM1, RIP1-mediated IKK complex recruitment / cellular response to amino acid stimulus / Downregulation of TGF-beta receptor signaling / EGFR downregulation / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Translesion synthesis by POLI / Gap-filling DNA repair synthesis and ligation in GG-NER / Josephin domain DUBs / Regulation of activated PAK-2p34 by proteasome mediated degradation / InlB-mediated entry of Listeria monocytogenes into host cell / IKK complex recruitment mediated by RIP1 Similarity search - Function
 Movie
Movie Controller
Controller

 Yorodumi
Yorodumi Open data
Open data
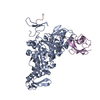
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information
 Homo sapiens (human)
Homo sapiens (human) X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads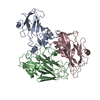








 PDBj
PDBj























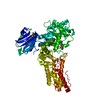
 Assembly
Assembly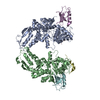


 Sample preparation
Sample preparation / Beamline: 24-ID-C / Wavelength: 0.9792
/ Beamline: 24-ID-C / Wavelength: 0.9792  Processing
Processing