| 登録情報 | データベース: PDB / ID: 4mmx
|
|---|
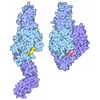
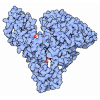
| タイトル | Integrin AlphaVBeta3 ectodomain bound to the tenth domain of Fibronectin |
|---|
 要素 要素 | - Fibronectin
 フィブロネクチン フィブロネクチン - Integrin alpha-VIntegrin alpha V
- Integrin beta-3Integrin beta 3
|
|---|
 キーワード キーワード | CELL ADHESION (細胞接着) / integrin (インテグリン) / A domain / hybrid domain / PSI / EGF repeats / beta TA thigh / beta propeller (Βプロペラドメイン) / RGD motif / fibronectin (フィブロネクチン) / vitronectin (ビトロネクチン) |
|---|
| 機能・相同性 |  機能・相同性情報 機能・相同性情報
integrin alphav-beta6 complex / integrin alphav-beta8 complex / transforming growth factor beta production / negative regulation of entry of bacterium into host cell / integrin alphav-beta5 complex / : / opsonin binding / negative regulation of monocyte activation / calcium-independent cell-matrix adhesion / integrin alphav-beta1 complex ...integrin alphav-beta6 complex / integrin alphav-beta8 complex / transforming growth factor beta production / negative regulation of entry of bacterium into host cell / integrin alphav-beta5 complex / : / opsonin binding / negative regulation of monocyte activation / calcium-independent cell-matrix adhesion / integrin alphav-beta1 complex / negative regulation of transforming growth factor beta production / Fibronectin matrix formation / Cross-presentation of particulate exogenous antigens (phagosomes) / extracellular matrix protein binding / Extracellular matrix organization / positive regulation of substrate-dependent cell migration, cell attachment to substrate / tube development / regulation of serotonin uptake / positive regulation of adenylate cyclase-inhibiting opioid receptor signaling pathway / alpha9-beta1 integrin-ADAM8 complex / regulation of trophoblast cell migration / neural crest cell migration involved in autonomic nervous system development / regulation of postsynaptic neurotransmitter receptor diffusion trapping / peptidase activator activity / alphav-beta3 integrin-vitronectin complex / maintenance of postsynaptic specialization structure / Laminin interactions / regulation of extracellular matrix organization / platelet alpha granule membrane / positive regulation of glomerular mesangial cell proliferation / fibrinogen complex / integrin alphav-beta3 complex / negative regulation of lipoprotein metabolic process / peptide cross-linking / alphav-beta3 integrin-PKCalpha complex / entry into host cell by a symbiont-containing vacuole / fibrinogen binding / glycinergic synapse / alphav-beta3 integrin-HMGB1 complex / integrin activation / vascular endothelial growth factor receptor 2 binding / blood coagulation, fibrin clot formation / negative regulation of lipid transport / ALK mutants bind TKIs / negative regulation of low-density lipoprotein receptor activity / Elastic fibre formation / regulation of phagocytosis / regulation of release of sequestered calcium ion into cytosol / cell-substrate junction assembly / mesodermal cell differentiation / transforming growth factor beta binding / angiogenesis involved in wound healing / alphav-beta3 integrin-IGF-1-IGF1R complex / platelet-derived growth factor receptor binding / positive regulation of small GTPase mediated signal transduction / filopodium membrane / extracellular matrix binding / positive regulation of fibroblast migration / positive regulation of vascular endothelial growth factor receptor signaling pathway / regulation of postsynaptic neurotransmitter receptor internalization / apolipoprotein A-I-mediated signaling pathway / regulation of bone resorption / wound healing, spreading of epidermal cells / apoptotic cell clearance / heterotypic cell-cell adhesion / positive regulation of cell adhesion mediated by integrin / integrin complex / biological process involved in interaction with symbiont / Molecules associated with elastic fibres / cellular response to insulin-like growth factor stimulus / proteoglycan binding / positive regulation of intracellular signal transduction / cell adhesion mediated by integrin / positive regulation of cell-matrix adhesion / smooth muscle cell migration / microvillus membrane / extracellular matrix structural constituent / MET activates PTK2 signaling / Syndecan interactions / negative chemotaxis / p130Cas linkage to MAPK signaling for integrins / cellular response to platelet-derived growth factor stimulus / cell-substrate adhesion / protein disulfide isomerase activity / positive regulation of smooth muscle cell migration / endodermal cell differentiation / activation of protein kinase activity / positive regulation of osteoblast proliferation / TGF-beta receptor signaling activates SMADs / PECAM1 interactions / lamellipodium membrane / GRB2:SOS provides linkage to MAPK signaling for Integrins / negative regulation of macrophage derived foam cell differentiation / negative regulation of lipid storage / platelet-derived growth factor receptor signaling pathway / fibronectin binding / Signaling by ALK fusions and activated point mutants / Non-integrin membrane-ECM interactions / 基底膜 / positive regulation of cell adhesion類似検索 - 分子機能 Hormone receptor fold - #30 / ntegrin, alpha v. Chain A, domain 4 / Integrin domains. Chain A, domain 2 / Hormone receptor fold / ntegrin, alpha v. Chain A, domain 3 / Integrin alpha, N-terminal / フィブロネクチンI型ドメイン / Fibronectin, type I / Fibronectin type-I domain signature. / Fibronectin type-I domain profile. ...Hormone receptor fold - #30 / ntegrin, alpha v. Chain A, domain 4 / Integrin domains. Chain A, domain 2 / Hormone receptor fold / ntegrin, alpha v. Chain A, domain 3 / Integrin alpha, N-terminal / フィブロネクチンI型ドメイン / Fibronectin, type I / Fibronectin type-I domain signature. / Fibronectin type-I domain profile. / Fibronectin type 1 domain / Integrin beta, epidermal growth factor-like domain 1 / Integrin beta epidermal growth factor like domain 1 / Integrin beta subunit, cytoplasmic domain / Integrin beta cytoplasmic domain / Integrin_b_cyt / : / Integrin alpha Ig-like domain 3 / Integrin beta tail domain / Integrin beta subunit, tail / Integrin beta tail domain superfamily / Integrin_B_tail / Integrin beta subunit, VWA domain / Integrin beta subunit / Integrin beta N-terminal / Integrin beta chain VWA domain / Integrin plexin domain / Integrins beta chain cysteine-rich domain signature. / Integrin beta subunits (N-terminal portion of extracellular region) / フィブロネクチンII型ドメイン / Fibronectin type II domain superfamily / フィブロネクチンII型ドメイン / Fibronectin type-II collagen-binding domain signature. / Fibronectin type-II collagen-binding domain profile. / Fibronectin type 2 domain / Integrin alpha cytoplasmic region / EGF-like domain, extracellular / EGF様ドメイン / Integrin alpha-2 / Integrin alpha Ig-like domain 1 / von Willebrand factor, type A domain / : / Integrin alpha Ig-like domain 2 / Integrin alpha chain / Integrin alpha beta-propellor / Integrin alpha chain, C-terminal cytoplasmic region, conserved site / Integrins alpha chain signature. / FG-GAP repeat profile. / Integrin alpha (beta-propellor repeats). / FG-GAP repeat / FG-GAP repeat / Integrin domain superfamily / Integrin alpha, N-terminal / PSI domain / domain found in Plexins, Semaphorins and Integrins / ラミニン / ラミニン / 7 Propeller / Methylamine Dehydrogenase; Chain H / Kringle-like fold / von Willebrand factor A-like domain superfamily / EGF-like domain signature 2. / EGF-like domain signature 1. / フィブロネクチンIII型ドメイン / Fibronectin type 3 domain / Fibronectin type-III domain profile. / Fibronectin type III / Fibronectin type III superfamily / リボン / Few Secondary Structures / イレギュラー / 抗体 / Immunoglobulin-like fold / Immunoglobulin-like / サンドイッチ / ロスマンフォールド / 3-Layer(aba) Sandwich / Mainly Beta / Alpha Beta類似検索 - ドメイン・相同性 : / フィブロネクチン / Integrin beta-3 / Integrin alpha-V類似検索 - 構成要素 |
|---|
| 生物種 |  Homo sapiens (ヒト) Homo sapiens (ヒト) |
|---|
| 手法 | X線回折 / シンクロトロン / 分子置換 / 解像度: 3.32 Å |
|---|
 データ登録者 データ登録者 | van Agthoven, J. / Xiong, J. / Arnaout, M.A. |
|---|
 引用 引用 | ジャーナル: Nat.Struct.Mol.Biol. / 年: 2014
タイトル: Structural basis for pure antagonism of integrin alpha V beta 3 by a high-affinity form of fibronectin.
著者: Van Agthoven, J.F. / Xiong, J.P. / Alonso, J.L. / Rui, X. / Adair, B.D. / Goodman, S.L. / Arnaout, M.A. |
|---|
| 履歴 | | 登録 | 2013年9月9日 | 登録サイト: RCSB / 処理サイト: RCSB |
|---|
| 改定 1.0 | 2014年3月26日 | Provider: repository / タイプ: Initial release |
|---|
| 改定 1.1 | 2014年4月9日 | Group: Database references |
|---|
| 改定 1.2 | 2014年4月30日 | Group: Database references |
|---|
| 改定 1.3 | 2017年11月15日 | Group: Refinement description / カテゴリ: software |
|---|
| 改定 2.0 | 2020年7月29日 | Group: Atomic model / Data collection ...Atomic model / Data collection / Database references / Derived calculations / Structure summary
カテゴリ: atom_site / chem_comp ...atom_site / chem_comp / entity / pdbx_branch_scheme / pdbx_chem_comp_identifier / pdbx_entity_branch / pdbx_entity_branch_descriptor / pdbx_entity_branch_link / pdbx_entity_branch_list / pdbx_entity_nonpoly / pdbx_nonpoly_scheme / pdbx_struct_assembly_gen / pdbx_struct_conn_angle / struct_asym / struct_conn / struct_ref_seq_dif / struct_site / struct_site_gen
Item: _atom_site.B_iso_or_equiv / _atom_site.Cartn_x ..._atom_site.B_iso_or_equiv / _atom_site.Cartn_x / _atom_site.Cartn_y / _atom_site.Cartn_z / _atom_site.auth_asym_id / _atom_site.auth_atom_id / _atom_site.auth_comp_id / _atom_site.auth_seq_id / _atom_site.label_asym_id / _atom_site.label_atom_id / _atom_site.label_comp_id / _atom_site.label_entity_id / _atom_site.type_symbol / _chem_comp.name / _chem_comp.type / _pdbx_struct_assembly_gen.asym_id_list / _pdbx_struct_conn_angle.ptnr1_auth_asym_id / _pdbx_struct_conn_angle.ptnr1_auth_comp_id / _pdbx_struct_conn_angle.ptnr1_auth_seq_id / _pdbx_struct_conn_angle.ptnr1_label_asym_id / _pdbx_struct_conn_angle.ptnr1_label_atom_id / _pdbx_struct_conn_angle.ptnr1_label_comp_id / _pdbx_struct_conn_angle.ptnr1_label_seq_id / _pdbx_struct_conn_angle.ptnr2_auth_asym_id / _pdbx_struct_conn_angle.ptnr2_auth_seq_id / _pdbx_struct_conn_angle.ptnr2_label_asym_id / _pdbx_struct_conn_angle.ptnr3_auth_asym_id / _pdbx_struct_conn_angle.ptnr3_auth_comp_id / _pdbx_struct_conn_angle.ptnr3_auth_seq_id / _pdbx_struct_conn_angle.ptnr3_label_asym_id / _pdbx_struct_conn_angle.ptnr3_label_atom_id / _pdbx_struct_conn_angle.ptnr3_label_comp_id / _pdbx_struct_conn_angle.ptnr3_label_seq_id / _pdbx_struct_conn_angle.value / _struct_conn.pdbx_dist_value / _struct_conn.pdbx_leaving_atom_flag / _struct_conn.pdbx_role / _struct_conn.ptnr1_auth_asym_id / _struct_conn.ptnr1_auth_comp_id / _struct_conn.ptnr1_auth_seq_id / _struct_conn.ptnr1_label_asym_id / _struct_conn.ptnr1_label_atom_id / _struct_conn.ptnr1_label_comp_id / _struct_conn.ptnr1_label_seq_id / _struct_conn.ptnr2_auth_asym_id / _struct_conn.ptnr2_auth_comp_id / _struct_conn.ptnr2_auth_seq_id / _struct_conn.ptnr2_label_asym_id / _struct_conn.ptnr2_label_atom_id / _struct_conn.ptnr2_label_comp_id / _struct_conn.ptnr2_label_seq_id / _struct_ref_seq_dif.details
解説: Carbohydrate remediation / Provider: repository / タイプ: Remediation |
|---|
|
|---|
 ムービー
ムービー コントローラー
コントローラー


 データを開く
データを開く
 基本情報
基本情報 構造の表示
構造の表示 ダウンロードとリンク
ダウンロードとリンク その他のダウンロード
その他のダウンロード










 PDBj
PDBj















 集合体
集合体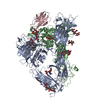



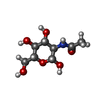
 タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 5 / 由来タイプ: 組換発現 / 式: C8H15NO6
タイプ: D-saccharide, beta linking / 分子量: 221.208 Da / 分子数: 5 / 由来タイプ: 組換発現 / 式: C8H15NO6

 分子量: 54.938 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Mn
分子量: 54.938 Da / 分子数: 8 / 由来タイプ: 合成 / 式: Mn 試料調製
試料調製 / ビームライン: 19-ID / 波長: 0.97921 Å
/ ビームライン: 19-ID / 波長: 0.97921 Å 解析
解析