 Movie
Movie Controller
Controller


+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9sfp | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Native cytoplasmic lattices from mouse oocytes | ||||||||||||||||||
 Components Components |
| ||||||||||||||||||
 Keywords Keywords | STRUCTURAL PROTEIN / oocyte / embryo / cytoskeleton / tubulin / ubiquitin | ||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationregulation of translation by machinery localization / Signaling by BMP / subcortical maternal complex / establishment of organelle localization / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Cilium Assembly / Sealing of the nuclear envelope (NE) by ESCRT-III / ZNF598 and the Ribosome-associated Quality Trigger (RQT) complex dissociate a ribosome stalled on a no-go mRNA / Chromatin modifying enzymes / protein storage ...regulation of translation by machinery localization / Signaling by BMP / subcortical maternal complex / establishment of organelle localization / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Cilium Assembly / Sealing of the nuclear envelope (NE) by ESCRT-III / ZNF598 and the Ribosome-associated Quality Trigger (RQT) complex dissociate a ribosome stalled on a no-go mRNA / Chromatin modifying enzymes / protein storage / structural constituent of cytoplasmic lattice / cytoplasmic lattice / cortical granule exocytosis / embryonic process involved in female pregnancy / ooplasm / endoplasmic reticulum localization / Intraflagellar transport / Carboxyterminal post-translational modifications of tubulin / establishment or maintenance of apical/basal cell polarity / COPI-independent Golgi-to-ER retrograde traffic / E3 ubiquitin ligases ubiquitinate target proteins / Downregulation of SMAD2/3:SMAD4 transcriptional activity / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / cytoplasm organization / histone H3K18 ubiquitin ligase activity / histone H3K14 ubiquitin ligase activity / histone H3 ubiquitin ligase activity / PINK1-PRKN Mediated Mitophagy / Inactivation of CSF3 (G-CSF) signaling / histone H3K23 ubiquitin ligase activity / spermatogonial cell division / COPI-mediated anterograde transport / Regulation of TNFR1 signaling / Kinesins / histone H3 reader activity / cortical granule / IKK complex recruitment mediated by RIP1 / PKR-mediated signaling / Aggrephagy / RHO GTPases activate IQGAPs / Oxygen-dependent proline hydroxylation of Hypoxia-inducible Factor Alpha / Mitotic Prometaphase / EML4 and NUDC in mitotic spindle formation / Resolution of Sister Chromatid Cohesion / Recycling pathway of L1 / The role of GTSE1 in G2/M progression after G2 checkpoint / COPI-dependent Golgi-to-ER retrograde traffic / positive regulation of embryonic development / apical cortex / chromosomal DNA methylation maintenance following DNA replication / positive regulation of meiotic nuclear division / regulation of RNA stability / axonemal microtubule / Hedgehog 'off' state / regulation of establishment of protein localization / RHO GTPases Activate Formins / Loss of Nlp from mitotic centrosomes / Recruitment of mitotic centrosome proteins and complexes / Loss of proteins required for interphase microtubule organization from the centrosome / organelle transport along microtubule / Anchoring of the basal body to the plasma membrane / Separation of Sister Chromatids / Recruitment of NuMA to mitotic centrosomes / AURKA Activation by TPX2 / hemi-methylated DNA-binding / forebrain morphogenesis / Peroxisomal protein import / glial cell differentiation / cerebellar cortex morphogenesis / Regulation of PLK1 Activity at G2/M Transition / dentate gyrus development / Neddylation / neuron projection arborization / regulation of epithelial cell proliferation / MHC class II antigen presentation / embryonic pattern specification / embryonic cleavage / mitochondrion localization / establishment of spindle localization / (E3-independent) E2 ubiquitin-conjugating enzyme / intermediate filament cytoskeleton / fertilization / Antigen processing: Ubiquitination & Proteasome degradation / methyl-CpG binding / protein K11-linked ubiquitination / protein K6-linked ubiquitination / pyramidal neuron differentiation / positive regulation of dendrite development / response to L-glutamate / epigenetic programming in the zygotic pronuclei / histone H3K9me2/3 reader activity / centrosome cycle / smoothened signaling pathway / 'de novo' protein folding / positive regulation of neurogenesis / E2 ubiquitin-conjugating enzyme / tubulin complex / negative regulation of gene expression via chromosomal CpG island methylation / mitotic spindle assembly checkpoint signaling / adult behavior Similarity search - Function | ||||||||||||||||||
| Biological species |  | ||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.2 Å | ||||||||||||||||||
 Authors Authors | Kilic, Z.I. / van Loenhout, J. / Chaillet, M. / Noteborn, W.E.M. / Leung, M.R. | ||||||||||||||||||
| Funding support |  Netherlands, 1items Netherlands, 1items
| ||||||||||||||||||
 Citation Citation | Journal: Nature / Year: 2026 Title: Cytoplasmic lattices are megadalton storage complexes in mammalian oocytes. Authors: Zeynep Ilgın Kılıç / Joyce van Loenhout / Marten Chaillet / Robert M van Es / Paula Sobrevals Alcaraz / Harmjan R Vos / Willem E M Noteborn / Miguel Ricardo Leung / Abstract: Mammalian oocytes store proteins for embryonic development on abundant structures called cytoplasmic lattices (CPLs). However, the mechanisms by which they achieve this are unclear, largely because ...Mammalian oocytes store proteins for embryonic development on abundant structures called cytoplasmic lattices (CPLs). However, the mechanisms by which they achieve this are unclear, largely because the molecular composition of the lattices themselves is unknown. Here we use cryo-electron microscopy and artificial intelligence-based modelling to reveal the molecular architecture and protein composition of native CPLs from mouse oocytes. We find that CPLs are formed by at least 13 different proteins that assemble into a megadalton-scale complex, including multiple copies of maternal effect factors such as PADI6 and the subcortical maternal complex. We show that proteins that are essential for early embryonic development are structural components of the CPLs, including the cytoskeletal proteins α-tubulin and β-tubulin, which are incorporated into CPLs as unpolymerized dimers, and an array of ubiquitination factors such as the epigenetic regulator and E3 ligase UHRF1, ubiquitin-conjugating E2 enzymes, and ubiquitin ligase substrate adapters. This represents an elegant molecular mechanism by which oocytes stockpile vital proteins through direct incorporation into highly stable supramolecular assemblies. Our findings provide a structural framework for understanding how disrupting stored maternal factors leads to infertility and developmental defects. | ||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9sfp.cif.gz | 2.5 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9sfp.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 9sfp.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/sf/9sfpftp://data.pdbj.org/pub/pdb/validation_reports/sf/9sfp | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  54838MC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj












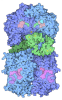





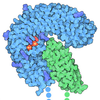












- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 | 
|
-Components
-Protein , 11 types, 24 molecules ABCDFKLMNOPQRSTUVWYabcde
| #1: Protein | Mass: 54036.707 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #2: Protein | | Mass: 48055.301 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #3: Protein | Mass: 108001.281 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #6: Protein | Mass: 18481.295 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #7: Protein | Mass: 76854.109 Da / Num. of mol.: 10 / Source method: isolated from a natural source / Source: (natural) #8: Protein | Mass: 65187.332 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #9: Protein | | Mass: 50188.441 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) References: UniProt: P68369, Hydrolases; Acting on acid anhydrides; Acting on GTP to facilitate cellular and subcellular movement #10: Protein | | Mass: 49877.824 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) #11: Protein | | Mass: 16706.133 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) References: UniProt: P61079, E2 ubiquitin-conjugating enzyme, (E3-independent) E2 ubiquitin-conjugating enzyme #12: Protein | | Mass: 88436.805 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) References: UniProt: Q8VDF2, RING-type E3 ubiquitin transferase #13: Protein | | Mass: 25643.373 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|
-NACHT, LRR and PYD domains-containing protein ... , 2 types, 3 molecules HIJ
| #4: Protein | Mass: 131468.547 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) #5: Protein | | Mass: 113527.188 Da / Num. of mol.: 1 / Source method: isolated from a natural source / Source: (natural) |
|---|
-Details
| Has protein modification | N |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: native cytoplasmic lattices from mouse oocytes / Type: COMPLEX / Entity ID: all / Source: NATURAL |
|---|---|
| Molecular weight | Experimental value: NO |
| Source (natural) | Organism: |
| Buffer solution | pH: 7.2 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Specimen support | Grid material: COPPER / Grid type: Quantifoil R2/1 |
| Vitrification | Cryogen name: ETHANE-PROPANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: TFS KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal magnification: 81000 X / Nominal defocus max: 2500 nm / Nominal defocus min: 500 nm / Cs: 2.7 mm / Alignment procedure: COMA FREE |
| Specimen holder | Cryogen: NITROGEN / Specimen holder model: FEI TITAN KRIOS AUTOGRID HOLDER |
| Image recording | Electron dose: 50 e/Å2 / Film or detector model: GATAN K3 BIOQUANTUM (6k x 4k) |
- Processing
Processing
| EM software |
| ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||||||||||||||
| Symmetry | Point symmetry: C2 (2 fold cyclic) | ||||||||||||||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.2 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 424595 / Symmetry type: POINT | ||||||||||||||||||||||||||||||||||||
| Atomic model building | Protocol: FLEXIBLE FIT |
