 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-9nh0: In situ cryo-EM structure of PR and DotA-IcmX of the Legionella D... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9nh0 | ||||||
|---|---|---|---|---|---|---|---|
| Title | In situ cryo-EM structure of PR and DotA-IcmX of the Legionella Dot/Icm T4SS machine at C1 symmetry | ||||||
 Components Components |
| ||||||
 Keywords Keywords | PROTEIN TRANSPORT / Type IVB Dot/Icm Secretion Machine | ||||||
| Function / homology |  Function and homology information Function and homology information | ||||||
| Biological species |  Legionella pneumophila subsp. pneumophila (bacteria) Legionella pneumophila subsp. pneumophila (bacteria) | ||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 4.63 Å | ||||||
 Authors Authors | Yue, J. / Liu, J. | ||||||
| Funding support |  United States, 1items United States, 1items
| ||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2025 Title: In situ structures of the Dot/Icm T4SS identify the DotA-IcmX complex as the gatekeeper for effector translocation. Authors: Jian Yue / Samira Heydari / Donghyun Park / David Chetrit / Shoichi Tachiyama / Wangbiao Guo / Jack M Botting / Shenping Wu / Craig R Roy / Jun Liu / Abstract: The Dot/Icm machine of is among the most versatile type IV secretion systems (T4SSs), capable of translocating more than 330 distinct effector proteins across the bacterial envelope into host cells. ...The Dot/Icm machine of is among the most versatile type IV secretion systems (T4SSs), capable of translocating more than 330 distinct effector proteins across the bacterial envelope into host cells. Assembly and function of the system require at least 27 Dot and Icm proteins, yet its architecture and activation mechanism remain poorly understood at the molecular level. Here, we deploy in situ single-particle cryoelectron microscopy to determine near-atomic structures of the Dot/Icm machine and its intimate association with three distinct outer membrane porins in intact bacteria. Notably, two essential yet enigmatic components, DotA and IcmX, form a pentameric protochannel in an inactive state at the central axis of the Dot/Icm machine. Upon Dot/Icm activation with host lysate, this protochannel undergoes extensive rearrangements to generate an extended transenvelope conduit, as visualized by cryoelectron tomography (cryo-ET) and subtomogram averaging. Furthermore, a combination of cryo-ET and cryo-FIB milling of macrophages infected with reveals tethering of the Dot/Icm machine to the host membrane, suggesting direct translocation of effector proteins from the bacterial cytoplasm into the host. Together, our studies identify the DotA-IcmX complex as a gatekeeper for effector translocation and provide a molecular framework for understanding the assembly and activation of the elaborate Dot/Icm T4SS. #1: Journal: bioRxiv / Year: 2025 Title: structures of the Dot/Icm T4SS identify the DotA-IcmX complex as the gatekeeper for effector translocation. Authors: Jian Yue / Samira Heydari / Donghyun Park / David Chetrit / Shoichi Tachiyama / Wangbiao Guo / Jack M Botting / Shenping Wu / Craig R Roy / Jun Liu Abstract: The Dot/Icm machine in is one of the most versatile type IV secretion systems (T4SSs), with a remarkable capacity to translocate over 330 different effector proteins across the bacterial envelope ...The Dot/Icm machine in is one of the most versatile type IV secretion systems (T4SSs), with a remarkable capacity to translocate over 330 different effector proteins across the bacterial envelope into host cells. At least 27 Dot and Icm proteins are required for assembly and function of the system, yet the architecture and activation mechanism remain poorly understood at the molecular level. Here, we deploy cryo-electron microscopy to reveal structures of the Dot/Icm machine at near-atomic resolution. Importantly, two proteins essential for effector translocation, DotA and IcmX, form a pentameric protochannel at an inactive state. Upon activation, the DotA-IcmX protochannel undergoes extensive rearrangements to form an extended transenvelope passage capable of transporting effector proteins from the bacterial cytoplasm into host cells as revealed by cryo-electron tomography. Collectively, our findings suggest that the DotA-IcmX complex functions as the gatekeeper for effector translocation of the Dot/Icm T4SS. | ||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9nh0.cif.gz | 2.8 MB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9nh0.ent.gz | Display | PDB format | |
| PDBx/mmJSON format | 9nh0.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/nh/9nh0ftp://data.pdbj.org/pub/pdb/validation_reports/nh/9nh0 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  49398MC  9nguC  9ngvC  9ngwC  9ngyC  9nh1C  9nh2C M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |





-Links
 PDBj
PDBj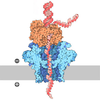


- Assembly
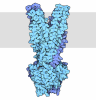
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Unknown peptide ... , 4 types, 72 molecules AaAgAmAsAyBeBkBqBwCcCiCoCuDaDgDmDsDyAbAhAnAtAzBfBlBrBxCdCjCp...
| #1: Protein/peptide | Mass: 1679.849 Da / Num. of mol.: 18 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria)#2: Protein/peptide | Mass: 910.095 Da / Num. of mol.: 18 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria)#3: Protein/peptide | Mass: 1775.978 Da / Num. of mol.: 18 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria)#4: Protein/peptide | Mass: 505.521 Da / Num. of mol.: 18 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria) |
|---|
-Protein , 5 types, 69 molecules AeAkAqAwBcBiBoBuCaCgCmCsCyDeDkDqDwEcAfAlArAxBdBjBpBvCbChCnCt...
| #5: Protein | Mass: 29729.969 Da / Num. of mol.: 18 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria)References: UniProt: Q5ZYC0 #6: Protein | Mass: 38958.926 Da / Num. of mol.: 18 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria)References: UniProt: Q5ZYC2 #7: Protein | Mass: 107911.891 Da / Num. of mol.: 23 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria)References: UniProt: Q5ZYC1 #8: Protein | Mass: 50712.422 Da / Num. of mol.: 5 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria)References: UniProt: Q5ZS30 #9: Protein | Mass: 113235.570 Da / Num. of mol.: 5 / Source method: isolated from a natural source Source: (natural) Legionella pneumophila subsp. pneumophila (bacteria)References: UniProt: Q5ZS33 |
|---|
-Details
| Has protein modification | Y |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: CELL / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: PR-protoChannel / Type: COMPLEX / Entity ID: all / Source: NATURAL |
|---|---|
| Molecular weight | Experimental value: NO |
| Source (natural) | Organism: Legionella pneumophila subsp. pneumophila (bacteria) |
| Buffer solution | pH: 6.7 |
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Microscopy | Model: FEI TECNAI 12 |
|---|---|
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 2000 nm / Nominal defocus min: 800 nm |
| Image recording | Electron dose: 73 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
- Processing
Processing
| EM software | Name: PHENIX / Version: 1.21.2_5419 / Category: model refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING ONLY | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 4.63 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 37503 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refinement | Cross valid method: NONE Stereochemistry target values: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| Displacement parameters | Biso mean: 260.1 Å2 | ||||||||||||||||||||||||
| Refine LS restraints |
|
