 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-9ne8: Human polymerase epsilon bound to PCNA and DNA with an in-situ-ge... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 9ne8 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Title | Human polymerase epsilon bound to PCNA and DNA with an in-situ-generated mismatch in the mismatch-locking state | |||||||||
 Components Components |
| |||||||||
 Keywords Keywords | REPLICATION / DNA polymerase / exo / holoenzyme / DNA | |||||||||
| Function / homology |  Function and homology information Function and homology informationDNA replication initiation / epsilon DNA polymerase complex / dinucleotide insertion or deletion binding / PCNA-p21 complex / mitotic telomere maintenance via semi-conservative replication / purine-specific mismatch base pair DNA N-glycosylase activity / nucleotide-excision repair, DNA gap filling / nuclear lamina / Polymerase switching / Processive synthesis on the lagging strand ...DNA replication initiation / epsilon DNA polymerase complex / dinucleotide insertion or deletion binding / PCNA-p21 complex / mitotic telomere maintenance via semi-conservative replication / purine-specific mismatch base pair DNA N-glycosylase activity / nucleotide-excision repair, DNA gap filling / nuclear lamina / Polymerase switching / Processive synthesis on the lagging strand / DNA replication proofreading / PCNA complex / single-stranded DNA 3'-5' DNA exonuclease activity / MutLalpha complex binding / Removal of the Flap Intermediate / Telomere C-strand (Lagging Strand) Synthesis / replisome / Mismatch repair (MMR) directed by MSH2:MSH3 (MutSbeta) / Mismatch repair (MMR) directed by MSH2:MSH6 (MutSalpha) / Transcription of E2F targets under negative control by DREAM complex / Polymerase switching on the C-strand of the telomere / mitotic DNA replication / response to L-glutamate / Processive synthesis on the C-strand of the telomere / Removal of the Flap Intermediate from the C-strand / Hydrolases; Acting on ester bonds; Exodeoxyribonucleases producing 5'-phosphomonoesters / response to dexamethasone / histone acetyltransferase binding / DNA synthesis involved in DNA repair / leading strand elongation / DNA polymerase processivity factor activity / G1/S-Specific Transcription / nuclear replication fork / SUMOylation of DNA replication proteins / replication fork processing / DNA repair-dependent chromatin remodeling / PCNA-Dependent Long Patch Base Excision Repair / embryonic organ development / Activation of the pre-replicative complex / response to cadmium ion / estrous cycle / mismatch repair / base-excision repair, gap-filling / cyclin-dependent protein kinase holoenzyme complex / DNA polymerase binding / translesion synthesis / epithelial cell differentiation / liver regeneration / TP53 Regulates Transcription of Genes Involved in G2 Cell Cycle Arrest / positive regulation of DNA replication / nuclear estrogen receptor binding / positive regulation of DNA repair / Translesion synthesis by REV1 / Translesion synthesis by POLK / Translesion synthesis by POLI / Gap-filling DNA repair synthesis and ligation in GG-NER / replication fork / G1/S transition of mitotic cell cycle / Termination of translesion DNA synthesis / Translesion Synthesis by POLH / receptor tyrosine kinase binding / Recognition of DNA damage by PCNA-containing replication complex / DNA-templated DNA replication / HDR through Homologous Recombination (HRR) / cellular response to hydrogen peroxide / Dual Incision in GG-NER / cellular response to UV / heart development / Dual incision in TC-NER / Gap-filling DNA repair synthesis and ligation in TC-NER / response to estradiol / mitotic cell cycle / E3 ubiquitin ligases ubiquitinate target proteins / 4 iron, 4 sulfur cluster binding / chromatin organization / DNA-directed DNA polymerase / damaged DNA binding / DNA-directed DNA polymerase activity / chromosome, telomeric region / DNA replication / nucleotide binding / centrosome / chromatin binding / chromatin / protein-containing complex binding / enzyme binding / DNA binding / extracellular exosome / nucleoplasm / zinc ion binding / identical protein binding / nucleus / plasma membrane Similarity search - Function | |||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human)synthetic construct (others) | |||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.6 Å | |||||||||
 Authors Authors | Wang, F. / He, Q. / Li, H. | |||||||||
| Funding support |  United States, 2items United States, 2items
| |||||||||
 Citation Citation | Journal: Proc Natl Acad Sci U S A / Year: 2025 Title: The proofreading mechanism of the human leading-strand DNA polymerase ε holoenzyme. Authors: Feng Wang / Qing He / Michael E O'Donnell / Huilin Li / Abstract: The eukaryotic leading-strand DNA polymerase ε (Polε) is a dual-function enzyme with a proofreading 3'-5' exonuclease () site located 40 Å from the DNA synthesizing site. Errors in Polε ...The eukaryotic leading-strand DNA polymerase ε (Polε) is a dual-function enzyme with a proofreading 3'-5' exonuclease () site located 40 Å from the DNA synthesizing site. Errors in Polε proofreading can cause various mutations, including C-to-G transversions, the most prevalent mutation in cancers and genetic diseases. Polε interacts with all three subunits of the PCNA ring to assemble a functional holoenzyme. Despite previous studies on proofreading of several Pol's, how Polε-or any Pol complexed with its sliding clamp-proofreads a mismatch generated in situ has been unknown. We show here by cryo-EM that a template/primer DNA substrate with a preexisting mismatch cannot enter the site of Polε-PCNA holoenzyme, but a mismatch generated in situ in the site yields three bona fide proofreading intermediates of Polε-PCNA holoenzyme. These intermediates reveal how the mismatch is dislodged from the site, how the DNA unwinds six base pairs, and how the unpaired primer 3'-end is inserted into the site for cleavage. These results unexpectedly demonstrate that PCNA imposes strong steric constraints that extend unwinding and direct the trajectory of mismatched DNA and that this trajectory is dramatically different than for Polε in the absence of PCNA. These findings suggest a physiologically relevant proofreading mechanism for the human Polε holoenzyme. | |||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 9ne8.cif.gz | 370.4 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb9ne8.ent.gz | 293.6 KB | Display | PDB format |
| PDBx/mmJSON format | 9ne8.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ne/9ne8ftp://data.pdbj.org/pub/pdb/validation_reports/ne/9ne8 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  49301MC  9ne6C  9ne7C  9ne9C  9neaC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |




-Links
 PDBj
PDBj








































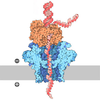







- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
| #1: Protein | Mass: 138137.562 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: POLE, POLE1Production host: Insect cell expression vector pTIE1 (others) References: UniProt: Q07864, DNA-directed DNA polymerase, Hydrolases; Acting on ester bonds; Exodeoxyribonucleases producing 5'-phosphomonoesters | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| #2: Protein | Mass: 28795.752 Da / Num. of mol.: 3 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PCNAProduction host:  References: UniProt: P12004 #3: DNA chain | | Mass: 10236.587 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) #4: DNA chain | | Mass: 14419.317 Da / Num. of mol.: 1 / Source method: obtained synthetically / Source: (synth.) synthetic construct (others) #5: Chemical | ChemComp-SF4 / | 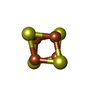  Mass: 351.640 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Fe4S4 / Feature type: SUBJECT OF INVESTIGATION Mass: 351.640 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: Fe4S4 / Feature type: SUBJECT OF INVESTIGATIONHas ligand of interest | Y | Has protein modification | N | |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: Ternary complex of human DNA polymerase epsilon with PCNA and the mismatched DNA Type: COMPLEX / Entity ID: #1-#4 / Source: RECOMBINANT | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Value: 0.22 MDa / Experimental value: NO | |||||||||||||||
| Source (natural) | Organism: Homo sapiens (human) | |||||||||||||||
| Source (recombinant) | Organism: Insect cell expression vector pTIE1 (others) | |||||||||||||||
| Buffer solution | pH: 7.5 | |||||||||||||||
| Buffer component |
| |||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | |||||||||||||||
| Vitrification | Instrument: FEI VITROBOT MARK IV / Cryogen name: ETHANE / Humidity: 100 % / Chamber temperature: 277 K |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: TFS KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 1600 nm / Nominal defocus min: 1200 nm |
| Image recording | Electron dose: 60 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
- Processing
Processing
| EM software | Name: PHENIX / Version: 1.21.2_5419 / Category: model refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: NONE | ||||||||||||||||||||||||
| Particle selection | Num. of particles selected: 15815155 | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.6 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 138854 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refinement | Highest resolution: 3.6 Å Stereochemistry target values: REAL-SPACE (WEIGHTED MAP SUM AT ATOM CENTERS) | ||||||||||||||||||||||||
| Refine LS restraints |
|
