 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 9f9s | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Yeast SDD1 Disome with Mbf1 | ||||||
 要素 要素 |
| ||||||
 キーワード キーワード | RIBOSOME / quality control / frame shifting / collision | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報GCN2-mediated signaling / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, LSU-rRNA,5S) / Negative regulators of DDX58/IFIH1 signaling / regulation of amino acid metabolic process / negative regulation of glucose mediated signaling pathway / positive regulation of translational fidelity / RMTs methylate histone arginines / Protein methylation / mTORC1-mediated signalling / Protein hydroxylation ...GCN2-mediated signaling / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, LSU-rRNA,5S) / Negative regulators of DDX58/IFIH1 signaling / regulation of amino acid metabolic process / negative regulation of glucose mediated signaling pathway / positive regulation of translational fidelity / RMTs methylate histone arginines / Protein methylation / mTORC1-mediated signalling / Protein hydroxylation / ribosome-associated ubiquitin-dependent protein catabolic process / GDP-dissociation inhibitor activity / positive regulation of nuclear-transcribed mRNA catabolic process, deadenylation-dependent decay / pre-mRNA 5'-splice site binding / Formation of the ternary complex, and subsequently, the 43S complex / Translation initiation complex formation / Ribosomal scanning and start codon recognition / preribosome, small subunit precursor / nonfunctional rRNA decay / response to cycloheximide / cleavage in ITS2 between 5.8S rRNA and LSU-rRNA of tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / Major pathway of rRNA processing in the nucleolus and cytosol / mRNA destabilization / SRP-dependent cotranslational protein targeting to membrane / GTP hydrolysis and joining of the 60S ribosomal subunit / negative regulation of translational frameshifting / Nonsense Mediated Decay (NMD) independent of the Exon Junction Complex (EJC) / Nonsense Mediated Decay (NMD) enhanced by the Exon Junction Complex (EJC) / negative regulation of mRNA splicing, via spliceosome / Formation of a pool of free 40S subunits / preribosome, large subunit precursor / L13a-mediated translational silencing of Ceruloplasmin expression / endonucleolytic cleavage to generate mature 3'-end of SSU-rRNA from (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translational elongation / ribosomal large subunit export from nucleus / G-protein alpha-subunit binding / 90S preribosome / positive regulation of protein kinase activity / Ub-specific processing proteases / ribosomal subunit export from nucleus / regulation of translational fidelity / protein-RNA complex assembly / endonucleolytic cleavage in ITS1 to separate SSU-rRNA from 5.8S rRNA and LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / translational termination / maturation of LSU-rRNA / ribosomal small subunit export from nucleus / translation regulator activity / DNA-(apurinic or apyrimidinic site) endonuclease activity / rescue of stalled ribosome / cellular response to amino acid starvation / ribosome assembly / maturation of LSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / ribosomal large subunit biogenesis / protein kinase C binding / maturation of SSU-rRNA from tricistronic rRNA transcript (SSU-rRNA, 5.8S rRNA, LSU-rRNA) / maturation of SSU-rRNA / translational initiation / small-subunit processome / macroautophagy / maintenance of translational fidelity / modification-dependent protein catabolic process / protein tag activity / cytoplasmic stress granule / rRNA processing / ribosome biogenesis / ribosome binding / ribosomal small subunit biogenesis / ribosomal small subunit assembly / small ribosomal subunit / 5S rRNA binding / ribosomal large subunit assembly / cytosolic small ribosomal subunit / large ribosomal subunit rRNA binding / small ribosomal subunit rRNA binding / cytosolic large ribosomal subunit / cytoplasmic translation / negative regulation of translation / rRNA binding / protein ubiquitination / ribosome / structural constituent of ribosome / G protein-coupled receptor signaling pathway / translation / negative regulation of gene expression / response to antibiotic / mRNA binding / ubiquitin protein ligase binding / nucleolus / mitochondrion / DNA binding / RNA binding / zinc ion binding / nucleoplasm / nucleus / cytosol / cytoplasm 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 2.9 Å | ||||||
 データ登録者 データ登録者 | Denk, T. / Beckmann, R. | ||||||
| 資金援助 |  ドイツ, 1件 ドイツ, 1件
| ||||||
 引用 引用 | ジャーナル: Mol Cell / 年: 2024 タイトル: Multiprotein bridging factor 1 is required for robust activation of the integrated stress response on collided ribosomes. 著者: Kyusik Q Kim / Jeffrey J Li / Ankanahalli N Nanjaraj Urs / Miguel E Pacheco / Victor Lasehinde / Timo Denk / Petr Tesina / Shota Tomomatsu / Yoshitaka Matsuo / Elesa McDonald / Roland ...著者: Kyusik Q Kim / Jeffrey J Li / Ankanahalli N Nanjaraj Urs / Miguel E Pacheco / Victor Lasehinde / Timo Denk / Petr Tesina / Shota Tomomatsu / Yoshitaka Matsuo / Elesa McDonald / Roland Beckmann / Toshifumi Inada / Rachel Green / Hani S Zaher /   要旨: In yeast, multiprotein bridging factor 1 (Mbf1) has been proposed to function in the integrated stress response (ISR) as a transcriptional coactivator by mediating a direct interaction between ...In yeast, multiprotein bridging factor 1 (Mbf1) has been proposed to function in the integrated stress response (ISR) as a transcriptional coactivator by mediating a direct interaction between general transcription machinery and the process's key effector, Gcn4. However, mounting evidence has demonstrated that Mbf1 (and its human homolog EDF1) is recruited to collided ribosomes, a known activator of the ISR. In this study, we connect these otherwise seemingly disparate functions of Mbf1. Our biochemical and structural analyses reveal that Mbf1 functions as a core ISR factor by interacting with collided ribosomes to mediate Gcn2 activation. We further show that Mbf1 serves no role as a transcriptional coactivator of Gcn4. Instead, Mbf1 is required for optimal stress-induced eukaryotic initiation factor 2α (eIF2α) phosphorylation and downstream de-repression of GCN4 translation. Collectively, our data establish that Mbf1 functions in ISR signaling by acting as a direct sensor of stress-induced ribosome collisions. #1: ジャーナル: Acta Crystallogr D Struct Biol / 年: 2019 タイトル: Macromolecular structure determination using X-rays, neutrons and electrons: recent developments in Phenix. 著者: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty / ...著者: Dorothee Liebschner / Pavel V Afonine / Matthew L Baker / Gábor Bunkóczi / Vincent B Chen / Tristan I Croll / Bradley Hintze / Li Wei Hung / Swati Jain / Airlie J McCoy / Nigel W Moriarty / Robert D Oeffner / Billy K Poon / Michael G Prisant / Randy J Read / Jane S Richardson / David C Richardson / Massimo D Sammito / Oleg V Sobolev / Duncan H Stockwell / Thomas C Terwilliger / Alexandre G Urzhumtsev / Lizbeth L Videau / Christopher J Williams / Paul D Adams /   要旨: Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological ...Diffraction (X-ray, neutron and electron) and electron cryo-microscopy are powerful methods to determine three-dimensional macromolecular structures, which are required to understand biological processes and to develop new therapeutics against diseases. The overall structure-solution workflow is similar for these techniques, but nuances exist because the properties of the reduced experimental data are different. Software tools for structure determination should therefore be tailored for each method. Phenix is a comprehensive software package for macromolecular structure determination that handles data from any of these techniques. Tasks performed with Phenix include data-quality assessment, map improvement, model building, the validation/rebuilding/refinement cycle and deposition. Each tool caters to the type of experimental data. The design of Phenix emphasizes the automation of procedures, where possible, to minimize repetitive and time-consuming manual tasks, while default parameters are chosen to encourage best practice. A graphical user interface provides access to many command-line features of Phenix and streamlines the transition between programs, project tracking and re-running of previous tasks. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 9f9s.cif.gz | 12.1 MB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb9f9s.ent.gz | 表示 | PDB形式 | |
| PDBx/mmJSON形式 | 9f9s.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/f9/9f9sftp://data.pdbj.org/pub/pdb/validation_reports/f9/9f9s | HTTPS FTP |
|---|
-関連構造データ



-リンク
 PDBj
PDBj






























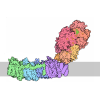



- 集合体
集合体
| 登録構造単位 | 
|
|---|---|
| 1 |
|
-要素
-タンパク質 , 6種, 9分子 CCCNLeMeSFRFSGRGMQ
| #1: タンパク質 | 分子量: 27272.611 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: SDD1, YEL057C / 発現宿主: | ||||||
|---|---|---|---|---|---|---|---|
| #3: タンパク質 | 分子量: 16434.682 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
| #27: タンパク質 | 分子量: 14583.077 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #55: タンパク質 | 分子量: 17254.227 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #56: タンパク質 | 分子量: 34841.219 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #89: タンパク質 | | 分子量: 33749.121 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-RNA鎖 , 4種, 4分子 CMCPDQDP
| #2: RNA鎖 | 分子量: 12722.189 Da / 分子数: 1 / 由来タイプ: 組換発現 由来: (組換発現) 発現宿主: |
|---|---|
| #4: RNA鎖 | 分子量: 24469.537 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #88: RNA鎖 | 分子量: 24801.801 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
| #91: RNA鎖 | 分子量: 24743.812 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
-Saccharomyces cerevisiae S288C ... , 5種, 8分子 L1L2M2L3M3S1R1M1
| #5: RNA鎖 | 分子量: 1097493.875 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) | ||||||
|---|---|---|---|---|---|---|---|
| #6: RNA鎖 | 分子量: 50682.922 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #7: RNA鎖 | 分子量: 38951.105 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #49: RNA鎖 | 分子量: 579761.938 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #87: RNA鎖 | | 分子量: 1098082.750 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) |
+60S ribosomal protein ... , 42種, 81分子 LALBMBLCMCLDMDLEMELFMFLGMGLHMHLIMILJMJLKMKLLMLLMMMLNMNLOMOLa...
-Small ribosomal subunit protein ... , 6種, 11分子 SARASCSDRDSdRdSuRuSvRv
| #50: タンパク質 | 分子量: 13538.922 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #52: タンパク質 | | 分子量: 7578.822 Da / 分子数: 1 / 由来タイプ: 天然 / 由来: (天然) #53: タンパク質 | 分子量: 6675.723 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #60: タンパク質 | 分子量: 26542.789 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #77: タンパク質 | 分子量: 13929.044 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) #78: タンパク質 | 分子量: 9758.829 Da / 分子数: 2 / 由来タイプ: 天然 / 由来: (天然) |
|---|
+40S ribosomal protein ... , 28種, 51分子 SBRBSERESaRaSbRbScRcSeReSfRfSgRgShRhSiRiSjRjSkRkSlRlSmRmSnRn...
-非ポリマー , 4種, 327分子 


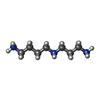
| #92: 化合物 | ChemComp-MG /  分子量: 24.305 Da / 分子数: 295 / 由来タイプ: 合成 / 式: Mg 分子量: 24.305 Da / 分子数: 295 / 由来タイプ: 合成 / 式: Mg#93: 化合物 | ChemComp-ZN /  分子量: 65.409 Da / 分子数: 13 / 由来タイプ: 合成 / 式: Zn 分子量: 65.409 Da / 分子数: 13 / 由来タイプ: 合成 / 式: Zn#94: 化合物 | ChemComp-K /  分子量: 39.098 Da / 分子数: 16 / 由来タイプ: 合成 / 式: K 分子量: 39.098 Da / 分子数: 16 / 由来タイプ: 合成 / 式: K#95: 化合物 |  分子量: 145.246 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C7H19N3 分子量: 145.246 Da / 分子数: 3 / 由来タイプ: 合成 / 式: C7H19N3 |
|---|
-詳細
| 研究の焦点であるリガンドがあるか | N |
|---|---|
| Has protein modification | N |
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 |
| ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 由来(天然) |
| ||||||||||||||||||||||||||||||
| 由来(組換発現) |
| ||||||||||||||||||||||||||||||
| 緩衝液 | pH: 7.5 | ||||||||||||||||||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | ||||||||||||||||||||||||||||||
| 急速凍結 | 凍結剤: ETHANE |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: SPOT SCAN |
| 電子レンズ | モード: BRIGHT FIELD / 最大 デフォーカス(公称値): 3500 nm / 最小 デフォーカス(公称値): 400 nm |
| 撮影 | 電子線照射量: 43.6 e/Å2 フィルム・検出器のモデル: GATAN K2 SUMMIT (4k x 4k) |
- 解析
解析
| EMソフトウェア | 名称: PHENIX / バージョン: 1.21.1_5286 / カテゴリ: モデル精密化 | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3次元再構成 | 解像度: 2.9 Å / 解像度の算出法: OTHER / 粒子像の数: 189911 詳細: This is a composite map. The masked resolution is given as calculated by Phenix. クラス平均像の数: 2 / 対称性のタイプ: POINT | ||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | ||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 74.43 Å2 | ||||||||||||||||||||||||
| 拘束条件 |
|
