XBP1(S) activates chaperone genes / target-directed miRNA degradation / elongin complex / reciprocal meiotic recombination / VCB complex / Cul5-RING ubiquitin ligase complex / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / Cul2-RING ubiquitin ligase complex / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery ...XBP1(S) activates chaperone genes / target-directed miRNA degradation / elongin complex / reciprocal meiotic recombination / VCB complex / Cul5-RING ubiquitin ligase complex / ubiquitin-dependent protein catabolic process via the C-end degron rule pathway / Cul2-RING ubiquitin ligase complex / Pausing and recovery of Tat-mediated HIV elongation / Tat-mediated HIV elongation arrest and recovery / HIV elongation arrest and recovery / Pausing and recovery of HIV elongation / Tat-mediated elongation of the HIV-1 transcript / Formation of HIV-1 elongation complex containing HIV-1 Tat / Formation of HIV elongation complex in the absence of HIV Tat / ubiquitin-like ligase-substrate adaptor activity / RNA Polymerase II Transcription Elongation / Formation of RNA Pol II elongation complex / RNA Polymerase II Pre-transcription Events / Maturation of protein E / Maturation of protein E / ER Quality Control Compartment (ERQC) / Myoclonic epilepsy of Lafora / FLT3 signaling by CBL mutants / IRAK2 mediated activation of TAK1 complex / Alpha-protein kinase 1 signaling pathway / Glycogen synthesis / IRAK1 recruits IKK complex / IRAK1 recruits IKK complex upon TLR7/8 or 9 stimulation / Prevention of phagosomal-lysosomal fusion / Endosomal Sorting Complex Required For Transport (ESCRT) / Membrane binding and targetting of GAG proteins / Regulation of TBK1, IKKε (IKBKE)-mediated activation of IRF3, IRF7 / Negative regulation of FLT3 / PTK6 Regulates RTKs and Their Effectors AKT1 and DOK1 / Regulation of TBK1, IKKε-mediated activation of IRF3, IRF7 upon TLR3 ligation / IRAK2 mediated activation of TAK1 complex upon TLR7/8 or 9 stimulation / Constitutive Signaling by NOTCH1 HD Domain Mutants / NOTCH2 Activation and Transmission of Signal to the Nucleus / TICAM1,TRAF6-dependent induction of TAK1 complex / TICAM1-dependent activation of IRF3/IRF7 / APC/C:Cdc20 mediated degradation of Cyclin B / Downregulation of ERBB4 signaling / APC-Cdc20 mediated degradation of Nek2A / Regulation of FZD by ubiquitination / p75NTR recruits signalling complexes / InlA-mediated entry of Listeria monocytogenes into host cells / TRAF6 mediated IRF7 activation in TLR7/8 or 9 signaling / NF-kB is activated and signals survival / TRAF6-mediated induction of TAK1 complex within TLR4 complex / Regulation of pyruvate metabolism / Pexophagy / Downregulation of ERBB2:ERBB3 signaling / Regulation of innate immune responses to cytosolic DNA / NRIF signals cell death from the nucleus / Regulation of PTEN localization / transcription corepressor binding / VLDLR internalisation and degradation / Activated NOTCH1 Transmits Signal to the Nucleus / Synthesis of active ubiquitin: roles of E1 and E2 enzymes / Translesion synthesis by REV1 / TICAM1, RIP1-mediated IKK complex recruitment / Regulation of BACH1 activity / Translesion synthesis by POLK / JNK (c-Jun kinases) phosphorylation and activation mediated by activated human TAK1 / InlB-mediated entry of Listeria monocytogenes into host cell / MAP3K8 (TPL2)-dependent MAPK1/3 activation / Activation of IRF3, IRF7 mediated by TBK1, IKKε (IKBKE) / Downregulation of TGF-beta receptor signaling / Translesion synthesis by POLI / Josephin domain DUBs / Gap-filling DNA repair synthesis and ligation in GG-NER / IKK complex recruitment mediated by RIP1 / PINK1-PRKN Mediated Mitophagy / TGF-beta receptor signaling in EMT (epithelial to mesenchymal transition) / TNFR1-induced NF-kappa-B signaling pathway / TP53 Regulates Transcription of DNA Repair Genes / Regulation of activated PAK-2p34 by proteasome mediated degradation / TCF dependent signaling in response to WNT / Regulation of NF-kappa B signaling / activated TAK1 mediates p38 MAPK activation / Autodegradation of Cdh1 by Cdh1:APC/C / APC/C:Cdc20 mediated degradation of Securin / NOTCH3 Activation and Transmission of Signal to the Nucleus / Regulation of signaling by CBL / Negative regulators of DDX58/IFIH1 signaling / N-glycan trimming in the ER and Calnexin/Calreticulin cycle / Asymmetric localization of PCP proteins / transcription initiation at RNA polymerase II promoter / Fanconi Anemia Pathway / Negative regulation of FGFR3 signaling / Ubiquitin-dependent degradation of Cyclin D / Peroxisomal protein import / Deactivation of the beta-catenin transactivating complex / SCF-beta-TrCP mediated degradation of Emi1 / NIK-->noncanonical NF-kB signaling / Stabilization of p53 / AUF1 (hnRNP D0) binds and destabilizes mRNA / TNFR2 non-canonical NF-kB pathway / Negative regulation of FGFR2 signaling Similarity search - Function
: / KLHDC2/KLHL20/DRC7 Kelch-repeats domain / Kelch motif / Kelch repeat type 1 / Elongin-C / Elongin B / Kelch-type beta propeller / S-phase kinase-associated protein 1-like / SKP1 component, POZ domain / Skp1 family, tetramerisation domain ...: / KLHDC2/KLHL20/DRC7 Kelch-repeats domain / Kelch motif / Kelch repeat type 1 / Elongin-C / Elongin B / Kelch-type beta propeller / S-phase kinase-associated protein 1-like / SKP1 component, POZ domain / Skp1 family, tetramerisation domain / Found in Skp1 protein family / SKP1/BTB/POZ domain superfamily / : / Ubiquitin domain signature. / Ubiquitin conserved site / Ubiquitin domain / Ubiquitin family / Ubiquitin homologues / Ubiquitin domain profile. / Ubiquitin-like domain / Ubiquitin-like domain superfamily Similarity search - Domain/homology
National Institutes of Health/National Institute on Aging (NIH/NIA)
AG011085
United States
Citation
Journal: Nat Commun / Year: 2024 Title: Structural basis for C-degron selectivity across KLHDCX family E3 ubiquitin ligases. Authors: Daniel C Scott / Sagar Chittori / Nicholas Purser / Moeko T King / Samuel A Maiwald / Kelly Churion / Amanda Nourse / Chan Lee / Joao A Paulo / Darcie J Miller / Stephen J Elledge / J Wade ...Authors: Daniel C Scott / Sagar Chittori / Nicholas Purser / Moeko T King / Samuel A Maiwald / Kelly Churion / Amanda Nourse / Chan Lee / Joao A Paulo / Darcie J Miller / Stephen J Elledge / J Wade Harper / Gary Kleiger / Brenda A Schulman / Abstract: Specificity of the ubiquitin-proteasome system depends on E3 ligase-substrate interactions. Many such pairings depend on E3 ligases binding to peptide-like sequences - termed N- or C-degrons - at the ...Specificity of the ubiquitin-proteasome system depends on E3 ligase-substrate interactions. Many such pairings depend on E3 ligases binding to peptide-like sequences - termed N- or C-degrons - at the termini of substrates. However, our knowledge of structural features distinguishing closely related C-degron substrate-E3 pairings is limited. Here, by systematically comparing ubiquitylation activities towards a suite of common model substrates, and defining interactions by biochemistry, crystallography, and cryo-EM, we reveal principles of C-degron recognition across the KLHDCX family of Cullin-RING ligases (CRLs). First, a motif common across these E3 ligases anchors a substrate's C-terminus. However, distinct locations of this C-terminus anchor motif in different blades of the KLHDC2, KLHDC3, and KLHDC10 β-propellers establishes distinct relative positioning and molecular environments for substrate C-termini. Second, our structural data show KLHDC3 has a pre-formed pocket establishing preference for an Arg or Gln preceding a C-terminal Gly, whereas conformational malleability contributes to KLHDC10's recognition of varying features adjacent to substrate C-termini. Finally, additional non-consensus interactions, mediated by C-degron binding grooves and/or by distal propeller surfaces and substrate globular domains, can substantially impact substrate binding and ubiquitylatability. Overall, the data reveal combinatorial mechanisms determining specificity and plasticity of substrate recognition by KLDCX-family C-degron E3 ligases.
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human) X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors United States, 1items
United States, 1items  Citation
Citation
 Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads




 PDBj
PDBj























 Assembly
Assembly
 Trichoplusia ni (cabbage looper) / References: UniProt: Q9BQ90
Trichoplusia ni (cabbage looper) / References: UniProt: Q9BQ90
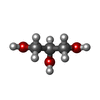

 Mass: 92.094 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C3H8O3 / Feature type: SUBJECT OF INVESTIGATION
Mass: 92.094 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C3H8O3 / Feature type: SUBJECT OF INVESTIGATION Sample preparation
Sample preparation Processing
Processing