Journal: Nat Commun / Year: 2024 Title: The EV71 2A protease occupies the central cleft of SETD3 and disrupts SETD3-actin interaction. Authors: Xiaopan Gao / Bei Wang / Kaixiang Zhu / Linyue Wang / Bo Qin / Kun Shang / Wei Ding / Jianwei Wang / Sheng Cui / Abstract: SETD3 is an essential host factor for the replication of a variety of enteroviruses that specifically interacts with viral protease 2A. However, the interaction between SETD3 and the 2A protease has ...SETD3 is an essential host factor for the replication of a variety of enteroviruses that specifically interacts with viral protease 2A. However, the interaction between SETD3 and the 2A protease has not been fully characterized. Here, we use X-ray crystallography and cryo-electron microscopy to determine the structures of SETD3 complexed with the 2A protease of EV71 to 3.5 Å and 3.1 Å resolution, respectively. We find that the 2A protease occupies the V-shaped central cleft of SETD3 through two discrete sites. The relative positions of the two proteins vary in the crystal and cryo-EM structures, showing dynamic binding. A biolayer interferometry assay shows that the EV71 2A protease outcompetes actin for SETD3 binding. We identify key 2A residues involved in SETD3 binding and demonstrate that 2A's ability to bind SETD3 correlates with EV71 production in cells. Coimmunoprecipitation experiments in EV71 infected and 2A expressing cells indicate that 2A interferes with the SETD3-actin complex, and the disruption of this complex reduces enterovirus replication. Together, these results reveal the molecular mechanism underlying the interplay between SETD3, actin, and viral 2A during virus replication.
Method to determine structure: MOLECULAR REPLACEMENT / Resolution: 3.52→19.99 Å / Cross valid method: THROUGHOUT / σ(F): 0 / Phase error: 31.23 / Stereochemistry target values: TWIN_LSQ_F Details: There are 4 pairs of molecules in the ASU. Only chains A/C have been conducted in-depth analysis and optimization and used for further structure analysis. Other molecules have poor electron ...Details: There are 4 pairs of molecules in the ASU. Only chains A/C have been conducted in-depth analysis and optimization and used for further structure analysis. Other molecules have poor electron density and were not carried out in-depth optimization.
Rfactor
Num. reflection
% reflection
Rfree
0.2914
2021
5.11 %
Rwork
0.2739
37546
-
obs
0.2762
39534
99.34 %
Solvent computation
Shrinkage radii: 0.9 Å / VDW probe radii: 1.11 Å / Solvent model: FLAT BULK SOLVENT MODEL
In the structure databanks used in Yorodumi, some data are registered as the other names, "COVID-19 virus" and "2019-nCoV". Here are the details of the virus and the list of structure data.
Jan 31, 2019. EMDB accession codes are about to change! (news from PDBe EMDB page)
EMDB accession codes are about to change! (news from PDBe EMDB page)
The allocation of 4 digits for EMDB accession codes will soon come to an end. Whilst these codes will remain in use, new EMDB accession codes will include an additional digit and will expand incrementally as the available range of codes is exhausted. The current 4-digit format prefixed with “EMD-” (i.e. EMD-XXXX) will advance to a 5-digit format (i.e. EMD-XXXXX), and so on. It is currently estimated that the 4-digit codes will be depleted around Spring 2019, at which point the 5-digit format will come into force.
The EM Navigator/Yorodumi systems omit the EMD- prefix.
Related info.:Q: What is EMD? / ID/Accession-code notation in Yorodumi/EM Navigator
Yorodumi is a browser for structure data from EMDB, PDB, SASBDB, etc.
This page is also the successor to EM Navigator detail page, and also detail information page/front-end page for Omokage search.
The word "yorodu" (or yorozu) is an old Japanese word meaning "ten thousand". "mi" (miru) is to see.
Related info.:EMDB / PDB / SASBDB / Comparison of 3 databanks / Yorodumi Search / Aug 31, 2016. New EM Navigator & Yorodumi / Yorodumi Papers / Jmol/JSmol / Function and homology information / Changes in new EM Navigator and Yorodumi
 Movie
Movie Controller
Controller


 Open data
Open data
 Basic information
Basic information Components
Components Keywords
Keywords Function and homology information
Function and homology information Homo sapiens (human)
Homo sapiens (human)
 Enterovirus A71
Enterovirus A71 X-RAY DIFFRACTION /
X-RAY DIFFRACTION /  Authors
Authors Citation
Citation
 Structure visualization
Structure visualization Downloads & links
Downloads & links Other downloads
Other downloads

 PDBj
PDBj
 Assembly
Assembly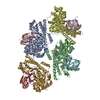




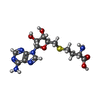
 Type: L-peptide linking / Mass: 384.411 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C14H20N6O5S / Feature type: SUBJECT OF INVESTIGATION
Type: L-peptide linking / Mass: 384.411 Da / Num. of mol.: 1 / Source method: obtained synthetically / Formula: C14H20N6O5S / Feature type: SUBJECT OF INVESTIGATION
 Mass: 65.409 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Zn / Feature type: SUBJECT OF INVESTIGATION
Mass: 65.409 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Zn / Feature type: SUBJECT OF INVESTIGATION Mass: 18.015 Da / Num. of mol.: 1417 / Source method: isolated from a natural source / Formula: H2O
Mass: 18.015 Da / Num. of mol.: 1417 / Source method: isolated from a natural source / Formula: H2O Sample preparation
Sample preparation Processing
Processing