 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8utp: KIF1A[1-393] - AMP-PNP two-heads-bound state in complex with a mi... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8utp | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | KIF1A[1-393] - AMP-PNP two-heads-bound state in complex with a microtubule - class T3L1 | |||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||
 Keywords Keywords | MOTOR PROTEIN / KIF1A / kinesin / motility / microtubule / tubulin | |||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationneuronal dense core vesicle membrane / dense core granule cytoskeletal transport / plus-end-directed kinesin ATPase / anterograde neuronal dense core vesicle transport / regulation of dendritic spine development / retrograde neuronal dense core vesicle transport / regulation of dendritic spine morphogenesis / anterograde axonal transport / plus-end-directed microtubule motor activity / Kinesins ...neuronal dense core vesicle membrane / dense core granule cytoskeletal transport / plus-end-directed kinesin ATPase / anterograde neuronal dense core vesicle transport / regulation of dendritic spine development / retrograde neuronal dense core vesicle transport / regulation of dendritic spine morphogenesis / anterograde axonal transport / plus-end-directed microtubule motor activity / Kinesins / Microtubule-dependent trafficking of connexons from Golgi to the plasma membrane / Resolution of Sister Chromatid Cohesion / Hedgehog 'off' state / Cilium Assembly / Intraflagellar transport / COPI-dependent Golgi-to-ER retrograde traffic / Mitotic Prometaphase / Carboxyterminal post-translational modifications of tubulin / RHOH GTPase cycle / EML4 and NUDC in mitotic spindle formation / Sealing of the nuclear envelope (NE) by ESCRT-III / Kinesins / PKR-mediated signaling / Separation of Sister Chromatids / The role of GTSE1 in G2/M progression after G2 checkpoint / Aggrephagy / RHO GTPases activate IQGAPs / RHO GTPases Activate Formins / HSP90 chaperone cycle for steroid hormone receptors (SHR) in the presence of ligand / MHC class II antigen presentation / Recruitment of NuMA to mitotic centrosomes / kinesin complex / COPI-mediated anterograde transport / COPI-dependent Golgi-to-ER retrograde traffic / cytoskeletal motor activity / neuronal dense core vesicle / microtubule-based process / vesicle-mediated transport / axon cytoplasm / microtubule cytoskeleton organization / structural constituent of cytoskeleton / microtubule cytoskeleton / mitotic cell cycle / microtubule binding / microtubule / Hydrolases; Acting on acid anhydrides; Acting on GTP to facilitate cellular and subcellular movement / axon / GTPase activity / synapse / dendrite / GTP binding / perinuclear region of cytoplasm / ATP hydrolysis activity / ATP binding / metal ion binding / identical protein binding / cytoplasm Similarity search - Function | |||||||||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / helical reconstruction / cryo EM / Resolution: 3.2 Å | |||||||||||||||||||||
 Authors Authors | Benoit, M.P.M.H. / Rao, L. / Asenjo, A.B. / Gennerich, A. / Sosa, H. | |||||||||||||||||||||
| Funding support |  United States, 6items United States, 6items
| |||||||||||||||||||||
 Citation Citation | Journal: Nat Commun / Year: 2024 Title: Cryo-EM unveils kinesin KIF1A's processivity mechanism and the impact of its pathogenic variant P305L. Authors: Matthieu P M H Benoit / Lu Rao / Ana B Asenjo / Arne Gennerich / Hernando Sosa / Abstract: Mutations in the microtubule-associated motor protein KIF1A lead to severe neurological conditions known as KIF1A-associated neurological disorders (KAND). Despite insights into its molecular ...Mutations in the microtubule-associated motor protein KIF1A lead to severe neurological conditions known as KIF1A-associated neurological disorders (KAND). Despite insights into its molecular mechanism, high-resolution structures of KIF1A-microtubule complexes remain undefined. Here, we present 2.7-3.5 Å resolution structures of dimeric microtubule-bound KIF1A, including the pathogenic P305L mutant, across various nucleotide states. Our structures reveal that KIF1A binds microtubules in one- and two-heads-bound configurations, with both heads exhibiting distinct conformations with tight inter-head connection. Notably, KIF1A's class-specific loop 12 (K-loop) forms electrostatic interactions with the C-terminal tails of both α- and β-tubulin. The P305L mutation does not disrupt these interactions but alters loop-12's conformation, impairing strong microtubule-binding. Structure-function analysis reveals the K-loop and head-head coordination as major determinants of KIF1A's superprocessive motility. Our findings advance the understanding of KIF1A's molecular mechanism and provide a basis for developing structure-guided therapeutics against KAND. | |||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8utp.cif.gz | 592.6 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8utp.ent.gz | 479.1 KB | Display | PDB format |
| PDBx/mmJSON format | 8utp.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ut/8utpftp://data.pdbj.org/pub/pdb/validation_reports/ut/8utp | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  42545MC  8utnC  8utoC  8utqC  8utrC  8utsC  8uttC  8utuC  8utvC  8utwC  8utyC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |










-Links
 PDBj
PDBj
















- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 3 types, 7 molecules KNAESBI
| #1: Protein | Mass: 49288.438 Da / Num. of mol.: 2 / Fragment: residues 1-393 Source method: isolated from a genetically manipulated source Details: "linker" residues are comprised of a leucine zipper based on S. cerevisiae GCN4, which is followed by the C-terminal strep-tag Source: (gene. exp.) Homo sapiens (human) / Gene: KIF1A, ATSV, C2orf20 / Production host:  #2: Protein | Mass: 50204.445 Da / Num. of mol.: 3 / Source method: isolated from a natural source / Source: (natural) #3: Protein | Mass: 49999.887 Da / Num. of mol.: 2 / Source method: isolated from a natural source / Source: (natural) |
|---|
-Non-polymers , 5 types, 13 molecules 
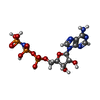
| #4: Chemical | ChemComp-MG /  Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg Mass: 24.305 Da / Num. of mol.: 4 / Source method: obtained synthetically / Formula: Mg#5: Chemical |  Mass: 506.196 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H17N6O12P3 / Comment: AMP-PNP, energy-carrying molecule analogue*YM Mass: 506.196 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H17N6O12P3 / Comment: AMP-PNP, energy-carrying molecule analogue*YM#6: Chemical |  Mass: 523.180 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM Mass: 523.180 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O14P3 / Comment: GTP, energy-carrying molecule*YM#7: Chemical |  Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM Type: RNA linking / Mass: 443.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O11P2 / Comment: GDP, energy-carrying molecule*YM#8: Chemical |  Mass: 853.906 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C47H51NO14 / Comment: medication, chemotherapy*YM Mass: 853.906 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C47H51NO14 / Comment: medication, chemotherapy*YM |
|---|
-Details
| Has ligand of interest | N |
|---|
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: FILAMENT / 3D reconstruction method: helical reconstruction |
- Sample preparation
Sample preparation
| Component | Name: KIF1A - AMP-PNP two-heads-bound state in complex with a microtubule - class T3LS Type: COMPLEX / Entity ID: #1-#3 / Source: MULTIPLE SOURCES | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular weight | Experimental value: NO | ||||||||||||||||||||||||
| Buffer solution | pH: 6.8 | ||||||||||||||||||||||||
| Buffer component |
| ||||||||||||||||||||||||
| Specimen | Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES | ||||||||||||||||||||||||
| Specimen support | Grid material: GOLD / Grid type: UltrAuFoil | ||||||||||||||||||||||||
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Experimental equipment |  Model: Titan Krios / Image courtesy: FEI Company |
|---|---|
| Microscopy | Model: FEI TITAN KRIOS |
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: BRIGHT FIELD / Nominal defocus max: 2000 nm / Nominal defocus min: 800 nm |
| Image recording | Electron dose: 50.22 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
- Processing
Processing
| Image processing |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Helical symmerty |
|