 Movie
Movie Controller
Controller

[English] 日本語
 Yorodumi
Yorodumi- PDB-8ui8: Cryo-EM map of human clamp-clamp loader ATAD5-RFC-two PCNAs compl... -
+ Open data
Open data

- Basic information
Basic information
| Entry | Database: PDB / ID: 8ui8 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Title | Cryo-EM map of human clamp-clamp loader ATAD5-RFC-two PCNAs complex in intermediate state 3 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Components Components |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Keywords Keywords | MOTOR PROTEIN / AAA ATPase / Clamp unloader | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Function / homology |  Function and homology information Function and homology informationDNA clamp unloader activity / positive regulation of cell cycle G2/M phase transition / positive regulation of DNA-directed DNA polymerase activity / Elg1 RFC-like complex / DNA replication factor C complex / Ctf18 RFC-like complex / nuclear DNA replication / dinucleotide insertion or deletion binding / PCNA-p21 complex / mitotic telomere maintenance via semi-conservative replication ...DNA clamp unloader activity / positive regulation of cell cycle G2/M phase transition / positive regulation of DNA-directed DNA polymerase activity / Elg1 RFC-like complex / DNA replication factor C complex / Ctf18 RFC-like complex / nuclear DNA replication / dinucleotide insertion or deletion binding / PCNA-p21 complex / mitotic telomere maintenance via semi-conservative replication / positive regulation of isotype switching to IgG isotypes / purine-specific mismatch base pair DNA N-glycosylase activity / DNA clamp loader activity / nuclear lamina / Polymerase switching / isotype switching / Processive synthesis on the lagging strand / DNA replication checkpoint signaling / PCNA complex / MutLalpha complex binding / Removal of the Flap Intermediate / Telomere C-strand (Lagging Strand) Synthesis / replisome / Mismatch repair (MMR) directed by MSH2:MSH3 (MutSbeta) / Mismatch repair (MMR) directed by MSH2:MSH6 (MutSalpha) / Transcription of E2F targets under negative control by DREAM complex / Polymerase switching on the C-strand of the telomere / mitotic DNA replication / response to L-glutamate / Processive synthesis on the C-strand of the telomere / Removal of the Flap Intermediate from the C-strand / negative regulation of intrinsic apoptotic signaling pathway in response to DNA damage by p53 class mediator / response to dexamethasone / HDR through Single Strand Annealing (SSA) / DNA strand elongation involved in DNA replication / histone acetyltransferase binding / regulation of mitotic cell cycle phase transition / DNA synthesis involved in DNA repair / leading strand elongation / DNA polymerase processivity factor activity / G1/S-Specific Transcription / Impaired BRCA2 binding to RAD51 / nuclear replication fork / SUMOylation of DNA replication proteins / intrinsic apoptotic signaling pathway in response to DNA damage by p53 class mediator / replication fork processing / DNA repair-dependent chromatin remodeling / PCNA-Dependent Long Patch Base Excision Repair / response to cadmium ion / Presynaptic phase of homologous DNA pairing and strand exchange / ATP-dependent activity, acting on DNA / Activation of ATR in response to replication stress / estrous cycle / mismatch repair / cyclin-dependent protein kinase holoenzyme complex / positive regulation of B cell proliferation / signal transduction in response to DNA damage / DNA polymerase binding / translesion synthesis / epithelial cell differentiation / liver regeneration / TP53 Regulates Transcription of Genes Involved in G2 Cell Cycle Arrest / positive regulation of DNA replication / nuclear estrogen receptor binding / positive regulation of DNA repair / Translesion synthesis by REV1 / Translesion synthesis by POLK / Translesion synthesis by POLI / Gap-filling DNA repair synthesis and ligation in GG-NER / replication fork / Termination of translesion DNA synthesis / Translesion Synthesis by POLH / receptor tyrosine kinase binding / Recognition of DNA damage by PCNA-containing replication complex / G2/M DNA damage checkpoint / cell population proliferation / DNA-templated DNA replication / HDR through Homologous Recombination (HRR) / cellular response to hydrogen peroxide / Dual Incision in GG-NER / cellular response to UV / heart development / Dual incision in TC-NER / Gap-filling DNA repair synthesis and ligation in TC-NER / response to estradiol / E3 ubiquitin ligases ubiquitinate target proteins / chromatin organization / Processing of DNA double-strand break ends / Regulation of TP53 Activity through Phosphorylation / damaged DNA binding / chromosome, telomeric region / DNA replication / DNA repair / centrosome / chromatin binding / chromatin / protein-containing complex binding / enzyme binding / ATP hydrolysis activity / DNA binding Similarity search - Function | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Biological species |  Homo sapiens (human) Homo sapiens (human) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Method | ELECTRON MICROSCOPY / single particle reconstruction / cryo EM / Resolution: 3.1 Å | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Authors Authors | Wang, F. / He, Q. / Li, H. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Funding support |  United States, 3items United States, 3items
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Citation Citation | Journal: Nat Struct Mol Biol / Year: 2024 Title: The human ATAD5 has evolved unique structural elements to function exclusively as a PCNA unloader. Authors: Feng Wang / Qing He / Nina Y Yao / Michael E O'Donnell / Huilin Li / Abstract: Humans have three different proliferating cell nuclear antigen (PCNA) clamp-loading complexes: RFC and CTF18-RFC load PCNA onto DNA, but ATAD5-RFC can only unload PCNA from DNA. The underlying ...Humans have three different proliferating cell nuclear antigen (PCNA) clamp-loading complexes: RFC and CTF18-RFC load PCNA onto DNA, but ATAD5-RFC can only unload PCNA from DNA. The underlying structural basis of ATAD5-RFC unloading is unknown. We show here that ATAD5 has two unique locking loops that appear to tie the complex into a rigid structure, and together with a domain that plugs the DNA-binding chamber, prevent conformation changes required for DNA binding, likely explaining why ATAD5-RFC is exclusively a PCNA unloader. These features are conserved in the yeast PCNA unloader Elg1-RFC. We observe intermediates in which PCNA bound to ATAD5-RFC exists as a closed planar ring, a cracked spiral or a gapped spiral. Surprisingly, ATAD5-RFC can open a PCNA gap between PCNA protomers 2 and 3, different from the PCNA protomers 1 and 3 gap observed in all previously characterized clamp loaders. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| History |
|
- Structure visualization
Structure visualization
| Structure viewer | Molecule: MolmilJmol/JSmol |
|---|
- Downloads & links
Downloads & links
-Download
| PDBx/mmCIF format | 8ui8.cif.gz | 513 KB | Display | PDBx/mmCIF format |
|---|---|---|---|---|
| PDB format | pdb8ui8.ent.gz | 402.4 KB | Display | PDB format |
| PDBx/mmJSON format | 8ui8.json.gz | Tree view | PDBx/mmJSON format | |
| Others |  Other downloads Other downloads |
-Validation report
| Arichive directory | https://data.pdbj.org/pub/pdb/validation_reports/ui/8ui8ftp://data.pdbj.org/pub/pdb/validation_reports/ui/8ui8 | HTTPS FTP |
|---|
-Related structure data
| Related structure data |  42288MC  8ui7C  8ui9C  8uiiC M: map data used to model this data C: citing same article ( |
|---|---|
| Similar structure data |
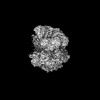
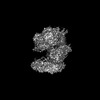
-Links
 PDBj
PDBj


















- Assembly
Assembly
| Deposited unit | 
|
|---|---|
| 1 |
|
-Components
-Protein , 2 types, 3 molecules AFG
| #1: Protein | Mass: 116421.094 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: ATAD5Production host: Insect cell expression vector pTIE1 (others) References: UniProt: Q96QE3 |
|---|---|
| #6: Protein | Mass: 28795.752 Da / Num. of mol.: 2 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: PCNAProduction host: Insect cell expression vector pTIE1 (others) References: UniProt: P12004 |
-Replication factor C subunit ... , 4 types, 4 molecules BCDE
| #2: Protein | Mass: 39203.207 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RFC2Production host: Insect cell expression vector pTIE1 (others) References: UniProt: P35250 |
|---|---|
| #3: Protein | Mass: 38545.512 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RFC5Production host: Insect cell expression vector pTIE1 (others) References: UniProt: P40937 |
| #4: Protein | Mass: 39735.711 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RFC4Production host: Insect cell expression vector pTIE1 (others) References: UniProt: P35249 |
| #5: Protein | Mass: 40614.332 Da / Num. of mol.: 1 Source method: isolated from a genetically manipulated source Source: (gene. exp.) Homo sapiens (human) / Gene: RFC3Production host: Insect cell expression vector pTIE1 (others) References: UniProt: P40938 |
-Non-polymers , 3 types, 8 molecules 
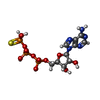

| #7: Chemical |  Mass: 24.305 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION Mass: 24.305 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: Mg / Feature type: SUBJECT OF INVESTIGATION#8: Chemical |  Mass: 523.247 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O12P3S / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP-gamma-S, energy-carrying molecule analogue*YM Mass: 523.247 Da / Num. of mol.: 3 / Source method: obtained synthetically / Formula: C10H16N5O12P3S / Feature type: SUBJECT OF INVESTIGATION / Comment: ATP-gamma-S, energy-carrying molecule analogue*YM#9: Chemical |  Mass: 427.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Feature type: SUBJECT OF INVESTIGATION / Comment: ADP, energy-carrying molecule*YM Mass: 427.201 Da / Num. of mol.: 2 / Source method: obtained synthetically / Formula: C10H15N5O10P2 / Feature type: SUBJECT OF INVESTIGATION / Comment: ADP, energy-carrying molecule*YM |
|---|
-Details
| Has ligand of interest | Y |
|---|---|
| Has protein modification | N |
-Experimental details
-Experiment
| Experiment | Method: ELECTRON MICROSCOPY |
|---|---|
| EM experiment | Aggregation state: PARTICLE / 3D reconstruction method: single particle reconstruction |
- Sample preparation
Sample preparation
| Component | Name: ATAD5-RFC-closed PCNA / Type: COMPLEX / Entity ID: #1-#6 / Source: RECOMBINANT |
|---|---|
| Molecular weight | Value: 0.32 MDa / Experimental value: NO |
| Source (natural) | Organism: Homo sapiens (human) |
| Source (recombinant) | Organism: Insect cell expression vector pTIE1 (others) |
| Buffer solution | pH: 7.5 |
| Specimen | Conc.: 1 mg/ml / Embedding applied: NO / Shadowing applied: NO / Staining applied: NO / Vitrification applied: YES |
| Vitrification | Cryogen name: ETHANE |
- Electron microscopy imaging
Electron microscopy imaging
| Microscopy | Model: FEI TITAN |
|---|---|
| Electron gun | Electron source:  FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM FIELD EMISSION GUN / Accelerating voltage: 300 kV / Illumination mode: FLOOD BEAM |
| Electron lens | Mode: DIFFRACTION / Nominal defocus max: 2000 nm / Nominal defocus min: 1000 nm |
| Image recording | Electron dose: 60 e/Å2 / Film or detector model: GATAN K3 (6k x 4k) |
- Processing
Processing
| EM software | Name: PHENIX / Category: model refinement | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF correction | Type: PHASE FLIPPING AND AMPLITUDE CORRECTION | ||||||||||||||||||||||||
| 3D reconstruction | Resolution: 3.1 Å / Resolution method: FSC 0.143 CUT-OFF / Num. of particles: 327653 / Symmetry type: POINT | ||||||||||||||||||||||||
| Refine LS restraints |
|
