 ムービー
ムービー コントローラー
コントローラー


+ データを開く
データを開く

- 基本情報
基本情報
| 登録情報 | データベース: PDB / ID: 8tdl | ||||||
|---|---|---|---|---|---|---|---|
| タイトル | Cryo-EM structure of the wild-type AtMSL10 in saposin | ||||||
 要素 要素 | Mechanosensitive ion channel protein 10 | ||||||
 キーワード キーワード | TRANSPORT PROTEIN / ion channels / mechanosensitive channels / heptamer / Arabidopsis thaliana | ||||||
| 機能・相同性 |  機能・相同性情報 機能・相同性情報programmed cell death in response to reactive oxygen species / leaf senescence / detection of mechanical stimulus / mechanosensitive monoatomic ion channel activity / monoatomic anion transport / plasma membrane 類似検索 - 分子機能 | ||||||
| 生物種 |  | ||||||
| 手法 | 電子顕微鏡法 / 単粒子再構成法 / クライオ電子顕微鏡法 / 解像度: 3.6 Å | ||||||
 データ登録者 データ登録者 | Zhang, J. / Yuan, P. | ||||||
| 資金援助 |  米国, 1件 米国, 1件
| ||||||
 引用 引用 | ジャーナル: Nat Commun / 年: 2023 タイトル: Open structure and gating of the Arabidopsis mechanosensitive ion channel MSL10. 著者: Jingying Zhang / Grigory Maksaev / Peng Yuan / 要旨: Plants are challenged by drastically different osmotic environments during growth and development. Adaptation to these environments often involves mechanosensitive ion channels that can detect and ...Plants are challenged by drastically different osmotic environments during growth and development. Adaptation to these environments often involves mechanosensitive ion channels that can detect and respond to mechanical force. In the model plant Arabidopsis thaliana, the mechanosensitive channel MSL10 plays a crucial role in hypo-osmotic shock adaptation and programmed cell death induction, but the molecular basis of channel function remains poorly understood. Here, we report a structural and electrophysiological analysis of MSL10. The cryo-electron microscopy structures reveal a distinct heptameric channel assembly. Structures of the wild-type channel in detergent and lipid environments, and in the absence of membrane tension, capture an open conformation. Furthermore, structural analysis of a non-conductive mutant channel demonstrates that reorientation of phenylalanine side chains alone, without main chain rearrangements, may generate the hydrophobic gate. Together, these results reveal a distinct gating mechanism and advance our understanding of mechanotransduction. | ||||||
| 履歴 |
|
- 構造の表示
構造の表示
| 構造ビューア | 分子: MolmilJmol/JSmol |
|---|
- ダウンロードとリンク
ダウンロードとリンク
-ダウンロード
| PDBx/mmCIF形式 | 8tdl.cif.gz | 738.6 KB | 表示 | PDBx/mmCIF形式 |
|---|---|---|---|---|
| PDB形式 | pdb8tdl.ent.gz | 581.5 KB | 表示 | PDB形式 |
| PDBx/mmJSON形式 | 8tdl.json.gz | ツリー表示 | PDBx/mmJSON形式 | |
| その他 |  その他のダウンロード その他のダウンロード |
-検証レポート
| 文書・要旨 | 8tdl_validation.pdf.gz | 1.2 MB | 表示 | wwPDB検証レポート |
|---|---|---|---|---|
| 文書・詳細版 | 8tdl_full_validation.pdf.gz | 1.2 MB | 表示 | |
| XML形式データ | 8tdl_validation.xml.gz | 65.7 KB | 表示 | |
| CIF形式データ | 8tdl_validation.cif.gz | 92.4 KB | 表示 | |
| アーカイブディレクトリ | https://data.pdbj.org/pub/pdb/validation_reports/td/8tdlftp://data.pdbj.org/pub/pdb/validation_reports/td/8tdl | HTTPS FTP |
-関連構造データ
| 関連構造データ |  41166MC  8tdjC  8tdkC  8tdmC M: このデータのモデリングに利用したマップデータ C: 同じ文献を引用 ( |
|---|---|
| 類似構造データ | |
| 実験データセット #1 | データ参照: 10.6019/EMPIAR-11612 / データの種類: EMPIAR / 詳細: EMPIAR-11612 |



-リンク
 PDBj
PDBj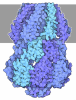

- 集合体
集合体
| 登録構造単位 | 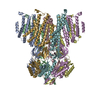
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 非結晶学的対称性 (NCS) | NCSドメイン:
NCSドメイン領域: Component-ID: 1 / Ens-ID: ens_1 / Beg auth comp-ID: THR / Beg label comp-ID: THR / End auth comp-ID: THR / End label comp-ID: THR / Auth seq-ID: 166 - 731 / Label seq-ID: 166 - 731
NCS oper:
|
-要素
| #1: タンパク質 | 分子量: 83955.562 Da / 分子数: 7 / 由来タイプ: 組換発現 由来: (組換発現) 遺伝子: MSL10 / 発現宿主:  Komagataella pastoris (菌類) / 参照: UniProt: Q9LYG9 Komagataella pastoris (菌類) / 参照: UniProt: Q9LYG9 |
|---|
-実験情報
-実験
| 実験 | 手法: 電子顕微鏡法 |
|---|---|
| EM実験 | 試料の集合状態: PARTICLE / 3次元再構成法: 単粒子再構成法 |
- 試料調製
試料調製
| 構成要素 | 名称: AtMSL10 in saposin / タイプ: COMPLEX / Entity ID: all / 由来: RECOMBINANT | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 分子量 | 実験値: NO | |||||||||||||||
| 由来(天然) | 生物種: | |||||||||||||||
| 由来(組換発現) | 生物種: Komagataella pastoris (菌類) | |||||||||||||||
| 緩衝液 | pH: 8 / 詳細: 20 mM Tris-HCl and 150 mM NaCl | |||||||||||||||
| 緩衝液成分 |
| |||||||||||||||
| 試料 | 包埋: NO / シャドウイング: NO / 染色: NO / 凍結: YES | |||||||||||||||
| 試料支持 | グリッドの材料: COPPER / グリッドのサイズ: 300 divisions/in. / グリッドのタイプ: Quantifoil R2/2 | |||||||||||||||
| 急速凍結 | 装置: FEI VITROBOT MARK IV / 凍結剤: ETHANE / 湿度: 100 % / 凍結前の試料温度: 277.15 K |
- 電子顕微鏡撮影
電子顕微鏡撮影
| 実験機器 |  モデル: Titan Krios / 画像提供: FEI Company |
|---|---|
| 顕微鏡 | モデル: FEI TITAN KRIOS |
| 電子銃 | 電子線源:  FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM FIELD EMISSION GUN / 加速電圧: 300 kV / 照射モード: FLOOD BEAM |
| 電子レンズ | モード: BRIGHT FIELD / 倍率(公称値): 59000 X / 最大 デフォーカス(公称値): 2400 nm / 最小 デフォーカス(公称値): 800 nm / Cs: 0.01 mm |
| 撮影 | 電子線照射量: 54 e/Å2 フィルム・検出器のモデル: FEI FALCON IV (4k x 4k) 実像数: 2120 |
- 解析
解析
| EMソフトウェア |
| |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTF補正 | タイプ: PHASE FLIPPING AND AMPLITUDE CORRECTION | |||||||||||||||||||||||||||||||||||||||||||||||||
| 粒子像の選択 | 選択した粒子像数: 404639 | |||||||||||||||||||||||||||||||||||||||||||||||||
| 対称性 | 点対称性: C7 (7回回転対称) | |||||||||||||||||||||||||||||||||||||||||||||||||
| 3次元再構成 | 解像度: 3.6 Å / 解像度の算出法: FSC 0.143 CUT-OFF / 粒子像の数: 69275 / 対称性のタイプ: POINT | |||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | プロトコル: RIGID BODY FIT | |||||||||||||||||||||||||||||||||||||||||||||||||
| 原子モデル構築 | PDB-ID: 8TDJ Accession code: 8TDJ / Source name: PDB / タイプ: experimental model | |||||||||||||||||||||||||||||||||||||||||||||||||
| 精密化 | 交差検証法: NONE 立体化学のターゲット値: GeoStd + Monomer Library + CDL v1.2 | |||||||||||||||||||||||||||||||||||||||||||||||||
| 原子変位パラメータ | Biso mean: 159.35 Å2 | |||||||||||||||||||||||||||||||||||||||||||||||||
| 拘束条件 |
| |||||||||||||||||||||||||||||||||||||||||||||||||
| Refine LS restraints NCS |
|
